


Chapter contents:
Arthropoda
–– 1. Stem-group Arthropods
–– 2. Trilobita ←
–– 3. Chelicerata
–– 4. Mandibulata
Fossil specimens of Eldredgeops crassituberculata from the Devonian of Ohio (USA). Image by James St. John; Creative Commons Attribution 2.0 Generic license.
Introduction
Trilobites are among the most well-known fossils, thanks in large part to their abundance, diversity, and broad distribution during the Paleozoic. These emblematic arthropods first appeared in the fossil record 521 million years ago and survived until the end-Permian extinction approximately 250 million years ago. Even at their first appearance, trilobites were diverse and widespread, suggesting a long history prior the first fossils, likely extending back earlier in time than the start of the Cambrian.
Trilobites are a well-studied group of organisms, yet their relationship to other arthropods is uncertain. Along with several other taxa with similar appearances, including the vicissicaudata that are introduced below, trilobites are often placed in the arthropod group Artiopoda. Depending on the analysis, artiopods have been grouped within the Mandibulata and Chelicerata, or as a clade of the same standing as these two groups. Until fossils from transitionary forms are found, the relationship of artiopods to other arthropods willl likely remain uncertain.
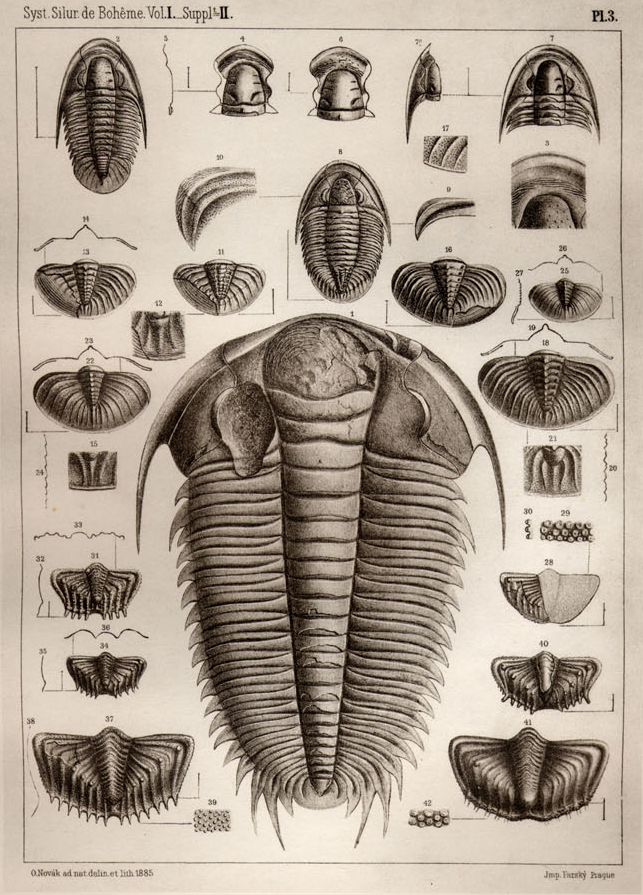
A selection of trilobites drawn by Joachim Barrande as a part of his early work on trilobites, published in 1852 (public domain).
{kind=link}
In part, the abundance of trilobites in the fossil record is due to their robust, hard skeleton and an arthropod characteristic: molting—also called ecdysis, which is the basis for the broader group name, Ecdysozoa, of which arthropods are a part. In addition to the remains of each trilobite when they died, their molts are also commonly found as fossils, enhancing their abundance in the record.
During the Cambrian, trilobites were a dominant component of marine ecosystems; however, many of the Cambrian trilobite species died out by the end of the Ordovician, leading up to and during the mass extinction at the end of this period. Though trilobite diversity decreased overall during this event (see image below), a portion of this diversity was offset by the evolution of new trilobite taxa. This second wave of trilobites diversified along with other marine organisms during the Great Ordovician Biodiversity Event, which was one of the most significant times of diversification that has ever occurred.
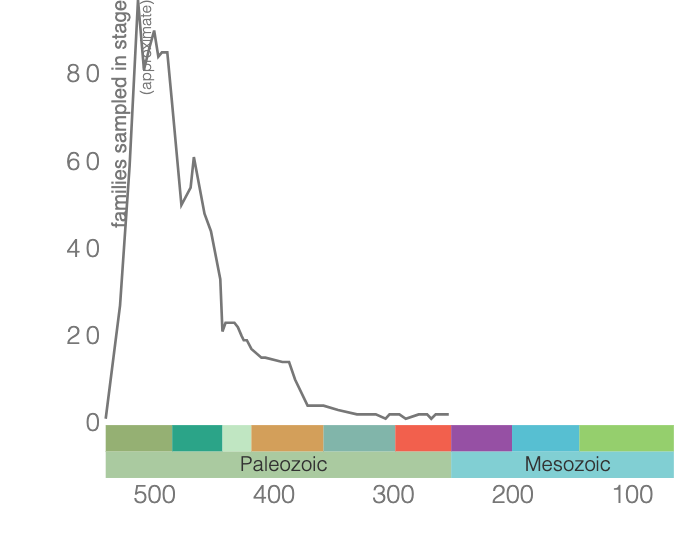
Family-level trilobite diversity. Image generated using the Paleobiology Database Navigator.
As increasingly powerful predators evolved during the early and middle Paleozoic, trilobites evolved defensive features to better protect themselves. These included morphological features like spines and behaviors like enrollment (see 3D model of Eldredgeops below). Even so, many fossil trilobites show evidence of bite marks from predators, which may have included eurypterids (learn more on the Stem-group Arthropod page) and early jawed fishes.
Fossil trilobite specimen of Eldredgeops crassituberculata from the Middle Devonian Silica Formation of Ohio (PRI 49811). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Longest dimension of specimen is approximately 5 cm. Model by Emily Hauf.
Starting in the Ordovician and for the remainder of the Paleozoic, family-level trilobite diversity declined until the extinction of this group during the end-Permian mass extinction. In the 270 million years trilobites were on Earth, more than 22,000 species existed, and more species continue to be discovered. Though the phylogenetic relationships of the different trilobite groups are still being determined, as is the placement of the Trilobita within the Arthropoda, trilobites remain useful to paleontologists for biostratigraphy. Because they evolved rapidly and were relatively widespread, trilobite species are often used to provide relative ages of the rocks in which they are found. You can explore some of this diversity in the 3D models, images, and videos on this page.
Watch "The Trouble with Trilobites," by PBS Eons (YouTube), to learn more about these Paleozoic arthropods.
Trilobite Anatomy
Like many other groups, the name trilobite is derived from the morphology of this taxon. Trilobite means “three lobes,” which refers to the longitudinal lobes that can be observed in most trilobites. As shown in the image below, they are called the left pleural lobe, axial lobe, and right pleural lobe.
Trilobites also exhibit the segmentation that is characteristic of all arthropods. In arthropods, these segments are called tagmata. The segmented body plan of arthropods, and trilobites, has played an important role in generating the diversity of species in this phylum. Simply put, the numerous segments, and variation within, provide a broad set of features on which natural selection can act. In general, trilobites have three tagmata, a cephalon, thorax, and pygidium (see the image below).
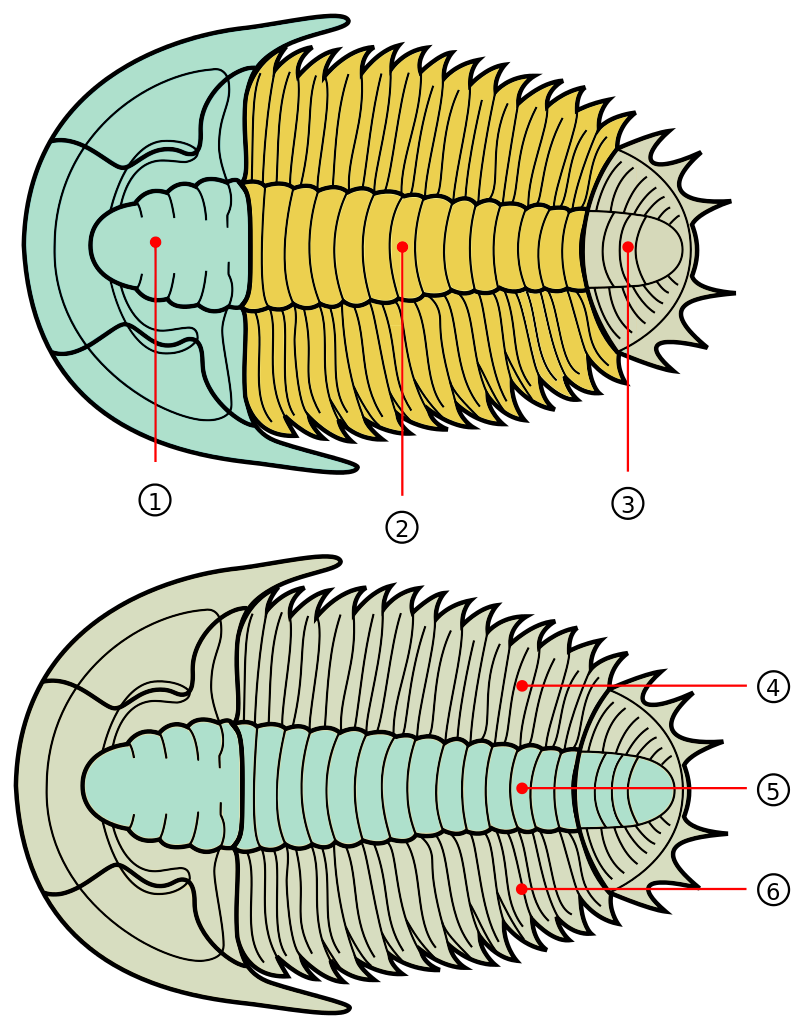
Trilobites have three major body sections, from anterior to posterios, called the cephalon (1), thorax (2), and pygidium (3). The body can also be divided into three longitudinal lobes, called the right pleural lobe (4), axial lobe (5), and left pleural lobe (6). Image by Sam Gon III; Creative Commons CC0 1.0 Universal Public Domain Dedication.
{kind=link}
Each segment also has constituent parts that can be variable between species. Considering the cephalon, or head segment, there are three general components: the glabella, fixigena, and librigena. The shape of these parts and the way in which they piece together—along sutures (internal black lines in the image below)—can also be highly variable. For example, the spines projecting backward in the librigenal can be absent, or accentuated, in some species. This flexibility in form led to the prolific diversity of the trilobites.
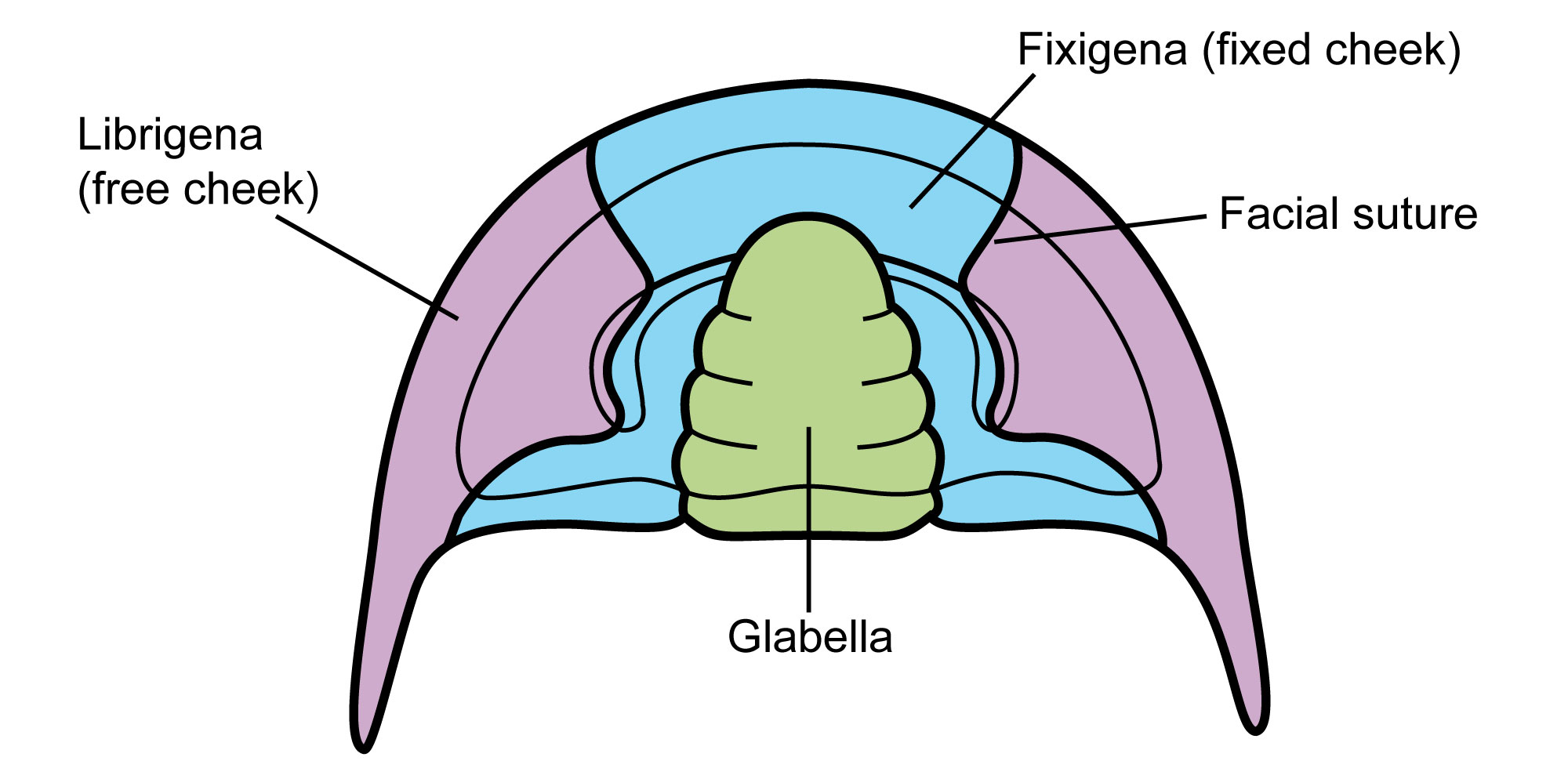
The head region (cephalon) of a trilobite contains a variety of elements, most notably the glabella, fixigena (or, fixed cheek), and librigena (or, free cheek). The facial suture separates the fixigena and librigena. The glabella and fixigena are collectively known as the cranidium. Image modified from original by Sam Gon III (pubilc domain).
{kind=link}
Like the cephalon, thorax morphology, particularly the number of repeated segments within the thorax, can vary considerably in different trilobite groups (explore the 3D models on this page for examples). The evolution of these segments and their differences between species reflect the different ecologies of the numerous trilobite species. As discussed in the video below, evolutionary modifications to the general trilobite body plan can tell paleontologists a great deal about how these animals lived.
Learn more about trilobite anatomy and how paleontologists use anatomy to interpret how these animals might have lived hundreds of millions of years ago. "Triumph of the Trilobites" by Ben G. Thomas (YouTube).
Occasionally, when preservational conditions were good, the soft-part anatomy of trilobites can also be studied (see panels A and B in the image below). In some cases, such as in the array of organisms in the image below, the tissues were replaced by pyrite (i.e., fools gold), contributing to their excellent preservation. These features, like segmented legs and antennae, provide additional information on the life mode of the trilobites and, by comparing these tissues between different organisms, can help determine phylogenetic relationships as well.

"New soft-bodied and delicate fossils from the new Beecher's-type beds. A: Ventral view of Triarthrus eatoni (Yale Peabody Museum, YPM 509231); note ostracod (arrowed, YPM 223943), like that in C, splayed across its midline. B: Lateral view of Triarthrus eatoni (YPM 223935). C: Ostracod (YPM 223936). Other notable fossils include the following. D: >30 legged ?crustacean (YPM 223937). E: Cryptolithus bellulus (YPM 509987). F: Ophiuroid (YPM 509229), and G: ?alga (YPM 223942). Scale bars = 1 cm, with exception of C, where scale bar = 1 mm. A, E, and G are from Martin Quarry, B and F are from Beecher's Trilobite Bed site, and C and D are from Jo's Quarry." Image and caption from Farrell et al. (2009) in Geology in accordance with Special "Fair Use" permission.
Explore trilobites and their anatomy in the 3D images below!
Fossil specimen of the trilobite Conocoryphe sulzeri from the Cambrian Jince Formation of the Czech Republic (PRI 76771). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Specimen (not including surrounding rock matrix) is approximately 5 cm in length. Model by Emily Hauf.
Fossil specimens of the trilobite Elrathia kingii from the Cambrian Wheeler Formation of Millard County, Utah (PRI 76837). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Length of larger specimen is about 4 cm; length of smaller specimen is about 3 cm. Model by Emily Hauf.
Fossil specimens of the trilobite Homotelus bromidensis from the Ordovician Bromide Formation of Carter County, Oklahoma (PRI 45505). Specimen from the collections of the Paleontological Research Institution, Ithaca, New York. Longest dimension of specimen is approximately 14 cm in length. Model by Emily Hauf.
Fossil specimen of the trilobite Flexicalymene meeki from the Ordovician of Cincinnati, Ohio (PRI 70754). Specimen is from the collections of the Paleontogical Research Institution, Ithaca, New York. Maximum diameter of specimen is approximately 2.5 cm. Model by Emily Hauf.
Fossil specimen of the trilobite Calymene celebra from the Silurian Niagara Series of Milwaukee, Wisconsin (PRI 42100). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Specimen is approximately 4 cm in length. Model by Emily Hauf.
Vicissicaudata
The Artiopoda, of which trilobites are a part, includes a variety of taxa, including Aglaspidida, Cheloniellida, Conciliterga, Nektaspidida, and Xandarellida), commonly grouped as the Trilobitomorpha and Vicissacaudata. These two groups are united by the presence of a post-abdominal segment lacking walking legs. As described by Lerosey-Aubril et al. (2017), the Vicissicaudata—which means “changed tailed” (see the tail-like structures in the images below)—include the aglaspidids (see the images below for examples), cheloniellids, and Cambrian the genera Emeraldella and Sidneyia. As with the trilobite phylogeny, the exact relationship between vicissicaudate taxa remains uncertain. Like trilobites, vicissicaudates were common in the Cambrian but dwindled in abundance through the Ordovician before going extinct.

An aglaspidid vicissicaudate, Cyclopites vulgaris, from the Cambrian of Wisconsin (USA). Image by Kennethcgass; Creative Commons Attribution-Share Alike 4.0 International license.
{kind=link}

An aglaspidid vicissicaudate, Beckwithia typa, from the Cambrian of Utah (USA). Image by Dwergenpaartje; Creative Commons Attribution-Share Alike 3.0 Unported license.
{kind=link}
References and Further Reading
Adrain, J. M., R. A. Fortey, and S. R. Westrop. 1998. Post-Cambrian trilobite diversity and evolutionary faunas. Science, 280: 1922-1925.
Eldredge, N. 1977. Trilobites and evolutionary patterns. Pp. 305-332, in: Developments in Palaeontology and Stratigraphy, A. Hallem (ed.), Elsevier, Amsterdam.
Farrell, Ú. C., M. J. Martin, J. W. Hagadorn, T. Whiteley, and D. E. Briggs. 2009. Beyond Beecher's Trilobite Bed: Widespread pyritization of soft tissues in the Late Ordovician Taconic foreland basin. Geology, 37: 907-910.
Fortey, R. A. 2001. Trilobite systematics: the last 75 years. Journal of Paleontology, 75: 1141-1151.
Lerosey-Aubril, R., X. Zhu, and J. Ortega-Hernández. 2017. The Vicissicaudata revisited–insights from a new aglaspidid arthropod with caudal appendages from the Furongian of China. Scientific Reports, 7: 1-18.
Lieberman, B. S. 1998. Cladistic analysis of the Early Cambrian olenelloid trilobites. Journal of Paleontology, 72: 59-78.
Lieberman, B. S., and N. Eldredge. 1996. Trilobite biogeography in the Middle Devonian: geological processes and analytical methods. Paleobiology, 22: 66-79.
Ortega‐Hernández, J., D. A. Legg, and S. J. Braddy. 2013. The phylogeny of aglaspidid arthropods and the internal relationships within Artiopoda. Cladistics, 29: 15-45.
Paterson, J. R. 2020. The trouble with trilobites: classification, phylogeny and the cryptogenesis problem. Geological Magazine, 157: 35-46.
Usage
Unless otherwise indicated, the written and visual content on this page is licensed under a Creative Commons Attribution-NonCommercial-Share Alike 4.0 International License. This page was written by Jansen A. Smith. See captions of individual images for attributions. See original source material for licenses associated with video and/or 3D model content.
