


Mollusca
– 2. Cephalopoda
–– 2.1 Cephalopoda stem groups
–– 2.2 "Nautiloidea"
–– 2.3 Ammonoidea
–– 2.4 Coleoidea ←
–– 2.5 Quiz
Above: A living chambered nautilus, photographed at the Melbourne Aquarium, Victoria, Australia. Photograph by Jonathan R. Hendricks.
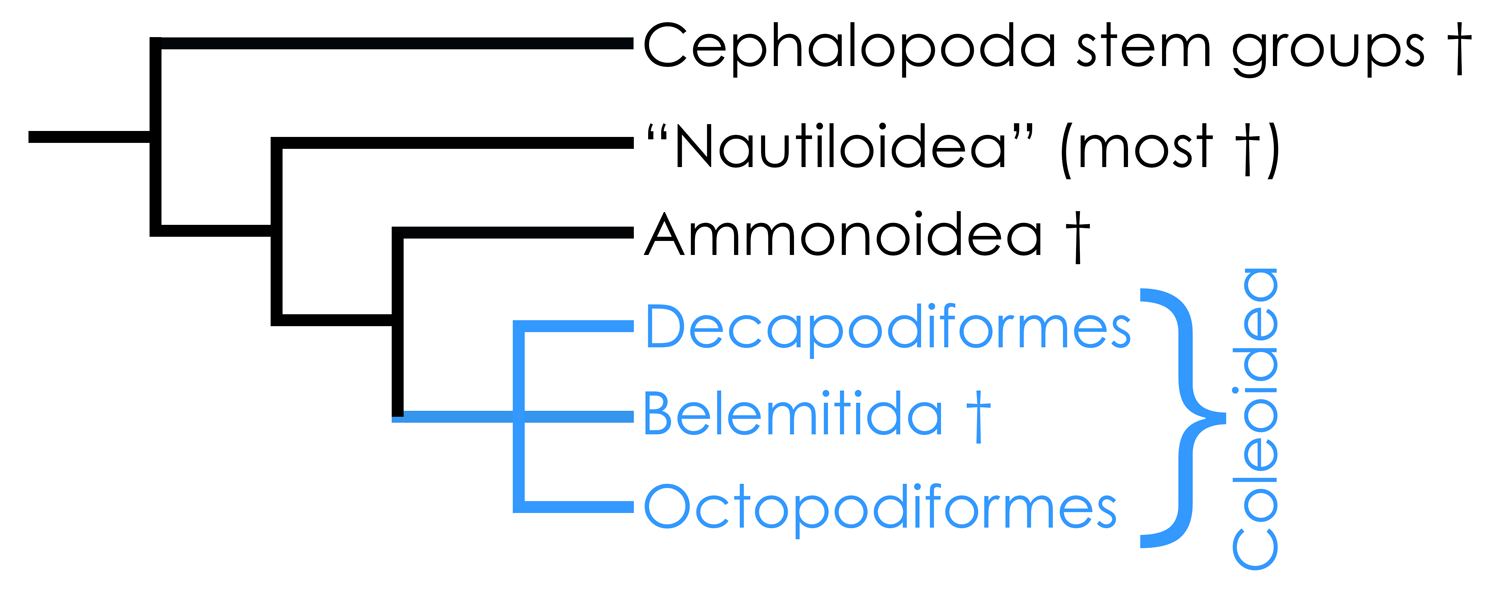
Highly simplified overview of cephalopod phylogeny based in part on the hypothesis of relationships presented by Kröger et al. (2011). Image by Jonathan R. Hendricks.
With the exception of the lone-surviving nautiloids (Nautilus and Allonautilus), all living cephalopods--including squids, cuttlefish, and octopuses--belong to the subclass Coleoidea, a truly remarkable animal clade. There are two major extant groups of coleoids (Decapodiformes and Octopodiformes), and one major extinct group (Belemnitida, the belemnites).
Coleoids differ significantly from nautiloids, ammonoids, and other extinct cephalopod groups in several respects. Perhaps most importantly, they lack external shells. In fact, many species do not have shells at all; the shells of those that do are completely internalized (though the argonaut, shown below, provides something of an exception). The story of how cephalopods went from a clade dominated by species with external shells to one nearly lacking them altogether is discussed in the video below.
Besides lacking an external shell, coleoids share a handful of other interesting attributes. For example, they all have the ability to rapidly change color using structures called chromatophores; they do this to better camouflage themselves, or to communicate with each other. The videos below provide some examples.
All species of coleoids also possess the ability to squirt ink to evade would-be predators.
Coleoid cephalopods also possess complex eyes that rival those of vertebrates in their visual capabilities (Gizmodo story).
Decapodiformes
Decapodiformes includes the squids and cuttlefish. As their name implies, all members of the group have ten arms (deca = ten; pod = foot), the forth pair of which are modified into tentacles that are often much longer than the other eight arms.
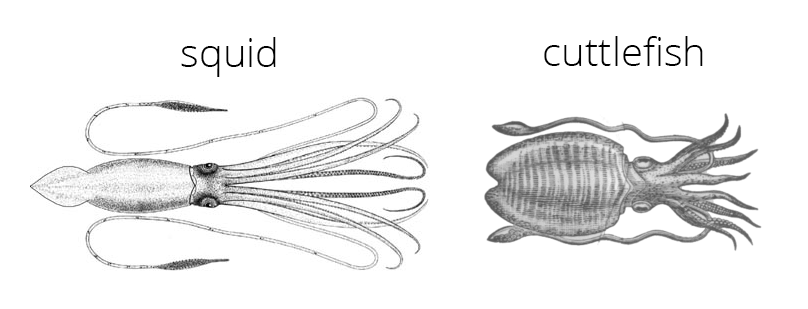
All Decapodiformes have ten arms, two of which are modified into long tentacles. Count them yourself in the images of the squid and cuttlefish above. Image sources: squid from Verrill (1879-1880; public domain); cuttlefish from Webster's Dictionary (1910; public domain).
Many modern squids possess a rigid internal structure called a gladius (plural: gladii; sometimes also called the pen) that is composed of chitin. Evidence suggests, however, that many ancient Octopodiformes also had gladii, which are absent in all modern octopods, except the vampire squid (see example below).
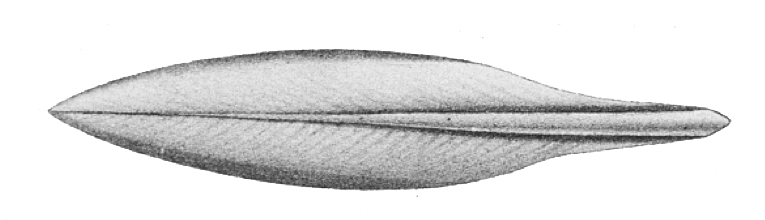
Drawing of the gladius of the extant squid Sepioteuthis lessoniana. Source: from L. Joubin (1900) (public domain).
{kind=link}
Cuttlefish possess a thin, chambered internal shell called a "cuttlebone" (see below).
Example 1: Colossal Squid
The colossal squid (Mesonychoteuthis hamiltoni) is the largest extant invertebrate animal, with some individuals exceeding 470 kg in mass (>1,000 lbs.) and 5.4 m in length (18 ft) (source: Museum of New Zealand).

A specimen of colossal squid (Mesonychoteuthis hamiltoni) collected from the Ross Sea, Antarctica. Image courtesy of the Museum of New Zealand (CC BY-NC-ND license).
The video below (which is a bit gross in places!) details several different features of the colossal squid, including its large beak (composed of chitin), huge eyes, and hooked tentacles.
Example 2: Humboldt Squid
Owing it its voracious feeding habits, the Humboldt squid (Dosidicus gigas) is sometimes also referred to as the "red devil." This species lives in the eastern Pacific and can grow to about 6 feet in length (1.8 m). The video below details its fierce nature.
Example 3: Spirula spirula
With the exception of Spirula spirula, modern squids lack internal shells. This deep-sea species possesses an internal, open-coiled shell that is used to maintain buoyancy. The animal itself swims in a vertical position, with the head pointed downward. For obvious reasons, the common name for this species is the ram's horn squid.

Drawing of Spirula spirula from Rübsamen (1910; public domain).

The internal shell of the squid Spirula spirula. The dark bands indicate the positions of internal septa that separate individual chambers. Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York (PRI 70572). Image by Jonathan R. Hendricks.
The very first video of Spirula spirula in its natural habitat was captured in October 2020 at a depth of over 2,700 feet near Australia's Great Barrier Reef. Surprisingly, the animal was discovered to orient itself with its head and tentacles pointing upwards, but its buoyant internal shell positioned downwards, the opposite of what had long been assumed.
Example 4: Color Changing Cuttlefishes
Cuttlefishes--a major subgroup of Decapodiformes--are able to very quickly change their coloration patterns to blend in with their habitats. This is well demonstrated in the video below.
Source: "Can cuttlefish camouflage in a living room? | Richard Hammond's Miracles of Nature" by BBC One.
Cuttlefishes have internal shells called "cuttlebones." These are composed of aragonite (calcium carbonate) and are sold in pet stores as a calcium source for parrots and other animals. The structure is internally chambered and, as in the chambered nautilus, is used by the cuttlefish to maintain buoyancy. A new septum is added daily in tropical species (Mangold and Young, 2016, Tree of Life).

A "cuttlebone," the internal, chambered shell of a cuttlefish (Sepia). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York (PRI 70571). Image by Jonathan R. Hendricks.
Fossil Record
While the internal shells of cuttlefish are known from the fossil record, there are no known definitive fossil squids that show preservation of the necessary ten arms. In fact, many fossil gladii previously attributed to "teuthids" (i.e., squids with gladii) are now thought by some scientists to belong to extinct species of gladius-bearing Octopodiformes. An interesting recent study by Clements et al. (2017) suggested that the complete absence of soft-bodied preservation of Decapodiformes may be attributed to the high amounts of ammonia in the living animals' bodies (used for buoyancy control), which renders soft-bodied preservation even less likely than normal.
Octopodiformes
Octopodiformes ("eight feet") includes the octopuses (also called octopods; despite being widely used, "octopi" is incorrect). There are nearly 300 extant species of octopuses (Gofas, 2009; WoRMS database accessed March 13, 2017). These eight-armed animals are the most intelligent invertebrates and anecdotal stories of their cleverness abound (examples: Scientific American, The Atlantic).
Some scientists alternatively use the name Vampyropoda for this group of coleoids, recognizing the membership of Vampyroteuthis infernalis--the vampire squid! (see Example 5 below)--in this clade.
Example 1: Giant Pacific Octopus
Octopuses--including the largest extant species, the giant Pacific octopus (Enteroctopus dofleini)--love to eat crabs. The video below demonstrates that they are clever animals capable of stealing crabs from traps left by fishermen.
Example 2: Mimic Octopus
As shown in the video below, the mimic octopus (Thaumoctopus mimicus) is remarkable for its ability to imitate the shapes and movements of other animals that live in its environment.
Example 3: Coconut Octopus
The video below shows a coconut octopus (Amphioctopus marginatus) that uses an empty bivalve shell as a portable lair; it also demonstrates the unusual manner by which this species sometimes "walks" about on two arms.
Example 4: The Argonaut (or, Paper Nautilus)
Female argonauts (genus Argonauta) look superficially like the chambered nautilus, but are actually octopuses. They secrete a very thin shell to hold their eggs. Unlike the nautilus, the external shell of the argonaut lacks chambers. The remarkable video below shows a passing group of argonauts.

The thin shell of Argonauta argo. Specimen collected from South Africa and is housed in the collections of the Paleontological Research Institution, Ithaca, new York (PRI 70575). Image by Jonathan R. Hendricks.
Example 5: The Vampire Squid
The vampire squid, Vampyroteuthis infernalis, is a bizarre coleoid that is the sole living representative of its taxonomic order, the Vampyromorpha. Its common name (the squid part, not the vampire part!) is suggestive of its confused taxonomic history: DNA evidence shows that it is more closely related to all other living octopuses than it is to squids (i.e., it is the "sister group" to other octopods). A recent study demonstrated--as shown in the video below--that this animal feeds on detritus suspended in seawater ("ocean snow") using a long filament that is extruded from its body, then reeled back in. This feeding mode is unique among extant cephalopods, as all other are active predators. The vampire squid is also remarkable for possessing a gladius, which is absent in all other octopods. This feature is important for interpreting gladii from the fossil record, however, as it supports the view that many of these fossils should be classified as Octopodiformes, rather than Decapodiformes (most especially since ten-armed coleoid fossils are unknown).
Fossil record
As might be predicted, the soft bodies of octopuses do not lend themselves to an extensive fossil record. That said, a modest number of species have been described from lagerstätten deposits. For example, a beautiful specimen of the octopod Proteroctopus ribeti from the Jurassic of France was recently imaged using synchroton microtomography by Kruta et al. (2016). It shows the characteristic "octopod" condition, as well as other combinations of characters that "indicates that Proteroctopus is a basal member of the Vampyropoda" (Kruta et al., 2016, p. 767). See Scientific American for a popular account of this amazing fossil, and BBC Earth for a short video about its significance.

Fossil specimen of the Jurassic octopod Proteroctopus ribeti from the La-Voulte-sur-Rhône lagerstatte of France. The roman numerals identify the four pairs of arms. Source: image published by Kruta et al. (2016) in Palaeontology; downloaded from MorphoBank (public domain).
As noted above, many modern squids possess rigid internal structures called gladii (singular: gladius; also called "pens"). Recent work summarized by Clements et al. (2017) suggests that many ancient Octopodiformes also had gladii, even though they are lacking in almost all modern species (an exception being the vampire squid; see above). Several such gladii from Cretaceous species from the Western Interior Seaway of North America are shown below.

Gladius of the species Euchoteuthis melanae, from the Cretaceous Niobrara Formation of Wichita County, Kansas. Specimen is from the collections of the Fort Hays State University's Sternberg Museum of Natural History (FHSMIP 1527). Image by Sternberg Museum of Natural History and Jonathan R. Hendricks

Gladius of species Kansasteuthis lindneri from the Cretaceous Niobrara Formation of Rooks County, Kansas. Specimen is from the collections of the Fort Hays State University's Sternberg Museum of Natural History (FHSMIP 1528). Image by Sternberg Museum of Natural History and Jonathan R. Hendricks

Gladius of Tusoteuthis longa from the Cretaceous Pierre Shale of Niobrara County, Wyoming. Specimen is from the collections of the Invertebrate Paleontology collections of the University of Kansas (KUMIP 150409). Image by KUMIP and Jonathan R. Hendricks
Belemnitida (Belemnites)
Belemnites were squid-like coleoid cephalopods that had an internal shell, the posterior portion of which--called the guard (or, rostrum)--was heavily calcified. These guards left behind a rich fossil record during the Jurassic and Cretaceous periods, though their simple, bullet-like shapes typically do not allow differentiation of species.

Guard of the belemnite Pachytheuthis densus from the Jurassic of Wyoming. Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York (PRI 70604). A 3D model of this same specimen is below. Image by Jonathan R. Hendricks.
Interactive 3D photogrammetry model of the belemnite Pachytheuthis densus from the Jurassic of Wyoming (PRI 70604). Length of specimen is approximately 11.5 cm. Model by Emily Hauf; Creative Commons Attribution-ShareAlike 4.0 International license (CC BY-SA 4.0).

Belemnites hastatus from the Jurassic of Germany, showing details of the internal shell. Specimen from the collections of the Paleontological Research Institution, Ithaca, New York (PRI 43506). Image by Jonathan R. Hendricks.
The specimen of Cylindroteuthis oweni shown in the interactive model below demonstrates how the guard surrounded the phragmocone in this particular species.
Interactive 3D photogrammetry model of the belemnite Cylindroteuthis oweni from the Jurassic Oxford Clay of Peterborough, England (Paleontological Research Institution, Ithaca, New York; PRI 70603). This specimen, which has an approximate length of 9.5 cm, shows a portion of the phragmocone nested inside of the guard. Model by Emily Hauf; Creative Commons Attribution-ShareAlike 4.0 International license (CC BY-SA 4.0).
While most of the fossil record of belemnites consists of guards, a modest number of fossils exhibiting soft-body preservation have been discovered. For example, the specimen below shows the distinctive calcitic guard on the right (posterior) side, ten arms bearing hooks on the left side, and a portion of the body (mantle) preserved in between.

An extraordinarily well-preserved specimen of belemnite, showing an intact guard, body, and ten arms bearing hooks. Source: photograph by "Ra'ike"; Creative Commons Attribution-Share Alike 3.0 Unported license.
{kind=link}
The specimen of Acanthoteuthis speciosa shown below only preserves the anterior end of the animal; numerous hook-bearing arms are evident.

Soft-part preservation showing the arms (with hooks) of the Jurassic belemnite Acanthoteuthis speciosa. This specimen from the Solnhofen Limestone of Germany and is housed in the collections of the Paleontological Research Institution, Ithaca, New York (PRI 70606). Image by Jonathan R. Hendricks.
References and further reading
Clements, T., C. Colleary, K. De Baets, and J. Vinther. 2017. Buoyancy mechanisms limit preservation of coleoid cephalopod soft tissues in mesozoic lagerstätten. Palaeontology 60(1): 1-14. Link.
Kröger, B., J. Vinther, and D. Fuchs. 2011. Cephalopod origin and evolution: a congruent picture emerging from fossils, development and molecules. Bioessays 33: 602-613. Link.
Kruta, I., I. Rouget, S. Charbonnier, J. Bardin, V. Fernandez, D. Germain, A. Brayard, and N. Landman. Proteroctopus ribeti in coleoid evolution. Palaeontology 59(6): 767-773. Link.
Staaf, D. 2017. Squid empire: the rise and fall of the cephalopods. ForeEdge, 256 pp.
Sutton, M. C. Perales-Raya, and I. Gilbert. 2016. A phylogeny of fossil and living neocoleoid cephalopods. Cladistics 32: 297-307. Link.
Tanner, A. R., D. Fuchs, I. E. Winkelmann, M. T. P. Gilbert, M. S. Pankey, Â. M. Ribeiro, K. M. Kocot, K. M. Halanych, T. H. Oakley, R. R. da Fonseca, D. Pisani, and J. Vinther. 2017. Molecular clocks indicate turnover and diversification of modern coleoid cephalopods during the Mesozoic Marine Revolution. Proceedings of the Royal Society B 284(1850): 1-7. Link.
Uribe, J. E. and R. Zardoya. 2017. Revisiting the phylogeny of Cephalopoda using complete mitochondrial genomes. Journal of Molluscan Studies 83: 133-144. Link.
Usage

Unless otherwise indicated, the written and visual content on this page is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. This page was written by Jonathan R. Hendricks. See captions of individual images for attributions. See original source material for licenses associated with video and/or 3D model content.