


Chapter contents:
Paleoecology
– 1. Paleoenvironmental Reconstruction ←
– 2. Biogeochemical Analysis and Paleoecology
– 3. Predation in Paleoecology
–– 3.1 Insect Herbivory in the Fossil Record
–– 3.2 Drilling Predation in the Fossil Record
–– 3.3 Dinosaur Predation in the Fossil Record
Above image by John Mason Clark (public domain).
{kind=link}
Introduction
In paleoecological studies, it is always important to understand the environment in which organisms were living. To understand this, think about your own surroundings as you go through a week in your life.
Is it warm, or cold? Is the weather more often rainy, or sunny? Are there other people around you? How about other species, like dogs, cats, trees, or insects?
The answers to these questions all help to describe your environment. When a paleoecologist studies an organism or community from the fossil record, these are the same types of questions they try to answer. Answering these questions about the past environment can provide important clues for understanding what we find in the fossil record.
Attempts to describe past environments are often referred to as paleoenvironmental reconstruction. The focus of paleoenvironmental reconstruction is often on abiotic factors like temperature, water depth, precipitation, or water salinity, but can also include biotic factors like the proportions of herbivores and predators or the number of species in a fossil assemblage. To reconstruct a paleoenvironment, information is gathered from a variety of sources and put together like clues in an investigation to reveal the setting in which organisms from a fossil assemblage interacted. These clues might include the type of sediments (e.g., sand, clay) that are present and features preserved in them (e.g., ripples, cross-bedding); taphonomic factors like the orientation of particles (i.e., indicating flow directions) and the degree to which fossils are disarticulated (i.e., energy in the system); climate data from tree rings, ice cores, or another source; and also the organisms in the assemblage, plus what we know about their biology. Once again, these interpretations rely on the assumption that processes and organisms functioned in the past just like they do today.

If you teleported and the first thing you saw was a palm tree, what type of environment would you think you landed in? What if the first thing you saw was a polar bear? Left image by Jawed Karim; Creative Commons Attribution-Share Alike 3.0 Unported license. Right image by Alan Wilson; Creative Commons Attribution-Share Alike 3.0 Unported license.
{kind=link}
{kind=link}
In many cases, there are not direct measurements of factors like rainfall and temperature and so paleoecologists need a different way to assess these factors. Often, this involves the use of a proxy. A proxy gives information about a factor, even if it does not directly measure something about that factor. Imagine that you develop the ability to teleport, but you have not quite figured out how to control where you are going. On your first attempt, you open your eyes and see palm trees blowing in the wind. What type of place have you arrived to? What if the first thing you saw was a polar bear?
Just like these organisms are associated with particular environments today, so are many fossil organisms. By using what we know about modern organisms and extending that knowledge to their long-dead relatives, we can make similar guesses about the type of environment that once existed. This type of information is referred to as a proxy. You did not measure the temperature, but when you saw a palm tree, you were able to guess that you were in a relatively warm environment. If you found a fossilized branch from a palm tree, you could reasonably draw the same conclusion. This type of proxy gives a relatively coarse indication of the past environment. You know it was warm, but you do not know exactly how warm it was in the past. Other types of proxies can give us different types of information and, when viewed together, give us a sense of the environment and ecosystem that once existed. We will explore a few more of these proxies.
Tree rings
Tree rings, the circular bands a tree grows each year as it gets bigger and bigger, are another good source of information about the past environment. Most studies of tree rings—referred to as a field as dendrochronology (dendro = relating to a tree; chronology = study of historical records to establish dates of past events)—occur using living trees. Scientists use a special borer to extract a small section of the tree, which allows them to examine the tree rings and does not kill the tree. Records like these can be used to go back thousands of years in time. Although these are not fossils, cores from living trees allow a dendrochronologist to correlate the information they see in the tree rings with information about temperature and rain fall that was recorded with modern instrumentation. For example, trees grow thicker rings when the climate is warm and wet and only grow thin rings during dry, cold times.

This ponderosa pine lived for more than 700 years, from 1255 - 1930, in Bly Oregon. If you look closely, you can see the years marked out on the stump. This section also shows damage from a fire, which led to the tree being cut down. Long term records like the rings of this tree provide high resolution information about the past environment. Image from Wikimedia Commons (public domain).
.jpg){kind=link}
This can happen seasonally (e.g., winter versus summer) or on longer timescales as climate changes over decades or centuries. These correlations allow dendrochronologists to be more confident in the conclusions they make using tree rings from times in the past when there were not measurements taken by instruments. The study of tree rings does not need to take place using living trees. Although it takes a little more work, scientists can use similar methods to study fossilized remains of trees to understand the temperature and rain fall occurring during the time those trees were alive. You can learn more about tree rings from NASA and NOAA.

A specimen of petrified wood (locality and age unknown) on display at the Natural History Museum, London. Note the preservation of tree rings, just as might be observed and counted on a modern log. Image by Jonathan R. Hendricks.  This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Ice cores
Ice cores, as with tree rings, can provide a high-resolution record of the past climate but have a somewhat limited duration of applicability, at least geologically speaking. This limitation is because in order to collect ice core data, there must be a continuous (or at least correlated) accumulation of ice, and there must be ice in the first place. Very few places on earth meet these requirements over timescales of centuries and millennia. Glaciers, at high elevations and altitudes, are one possibility for data collection, but the best ice core data come from the polar ice sheets of Greenland and Antarctica. In these cold, polar environments, snow and ice have been accumulating for 130,000 and 800,000 years, respectively! The core extracted from Antarctica, which was over two miles long, provides the best record we have for climate during the last million years and, because it includes layers from more recent times when modern instruments were recording climate, we can have strong confidence in the evidence it gives us.

"This photograph shows a section of the GISP2 ice core from 1837-1838 meters in which annual layers are clearly visible. The appearance of layers results from differences in the size of snow crystals deposited in winter versus summer and resulting variations in the abundance and size of air bubbles trapped in the ice. Counting such layers has been used (in combination with other techniques) to reliably determine the age of the ice. This ice was formed ~16250 years ago during the final stages of the last ice age and approximately 38 years are represented here. By analyzing the ice and the gases trapped within, scientists are able to learn about past climate conditions." Image and caption from by USGS staff (Wikimedia Commons; public domain).
{kind=link}
The high resolution of ice core data is due to the yearly accumulation of snow, ice, and gas bubbles trapped inside. It’s not always easy to see in the core, but if you look closely at the image above, you can clearly see some of these annual bands. Over time, the growing weight of accumulating ice compacts the layers below and preserves the climate information in a deep-freeze. Importantly, the ice and snow also collect things like dust, ash, pollen, salts, and anything else that was in the atmosphere when the layer formed. Analysis of these particles can provide a wealth of information about the past climate. Most notably, the Antarctica ice core allows us to reconstruct the amounts of greenhouse gases that were in the atmosphere during the last several hundred thousand years. These data show clear evidence of the climate change we are experiencing today—compare today’s carbon dioxide concentration of more than 410 ppm with concentrations from the past (blue lines).
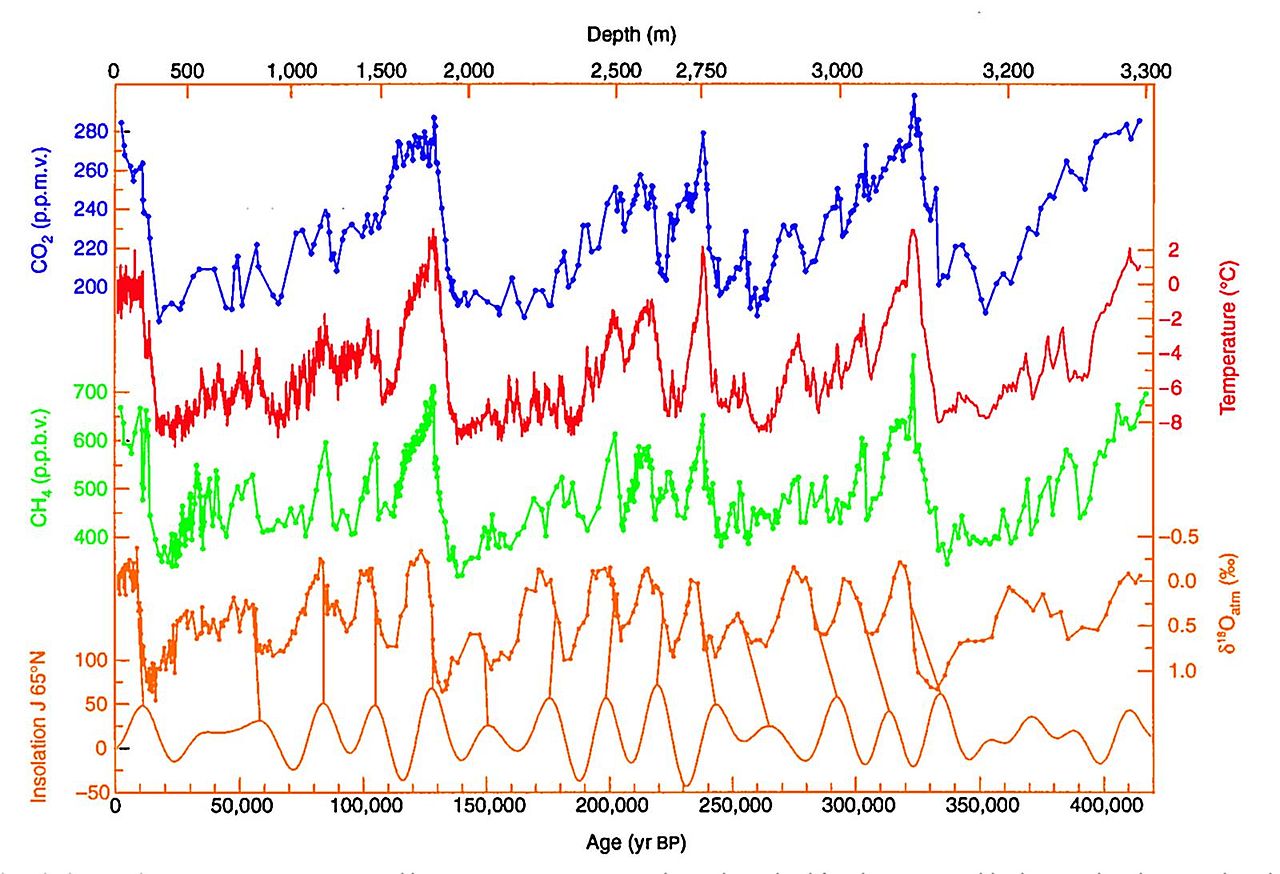
This graph shows the atmospheric concentrations of carbon dioxide (top on left x-axis, blue) and methane (middle on left x-axis, green) over the last 400,000 years, as indicated by analysis of ice core samples. Over this time, the graph also shows reconstructed temperature (top on right x-axis, red), the ratio of 18O:16O (bottom on right x-axis, orange), and amount of energy received from the sun (bottom on left x-axis, orange). These high resolution data show a natural cycle in the past, as concentrations of carbon dioxide and methane in the atmosphere drove warming and cooling cycles over 100,000 years periods. Image from Wikimedia Commons (public domain).
{kind=link}
You can learn more about ice cores, including how they are sampled, stored, and analyzed from the National Science Foundation Ice Core Facility. The video below will give you a brief introduction to the work they do at the Ice Core Facility.
"Ancient Ice Reveals Earth's Climate History" by Associated Press. Source: YouTube.
Sediments and their features
Ice cores and tree rings can provide high resolution, relatively recent data on the past environment. Particularly when studying life in the last thousands to million years, these data can be incredibly useful. When we go further back in time, however, other proxies such as sediment type, sediment features, taphonomic processes, and organismal biology play a larger role in reconstructing the paleoenvironment.
Each piece of information, like the degree of disarticulation, orientation of particles, or types of sediments, helps to develop a more complete picture of the past environment. Each piece on its own may not be sufficient to confidently characterize the past environment but, taken together as multiple lines of evidence, they can tell a compelling story.
Sediments
The sediments in which a fossil assemblage is preserved can provide information about the type of environment where the organisms lived, especially for marine ecosystems. Sediments (physical matter found in the natural world and subject to processes like weathering and erosion) come in all sorts of shapes and sizes, ranging from huge boulders to tiny flecks of clay. Based on how big or small these particles are, a paleoecologist can get a good sense of the amount of energy in the system and using this can pick up clues about the paleoenvironment.

Classification of sedimentary grains, which is based on grain size. Image by Jonathan R. Hendricks.
To understand this concept, imagine that you need to pick up two pieces of sediment, a boulder with a diameter of one foot and a single grain of sand. Which of these could you carry for a longer distance? No matter who you are and how strong you might be, you can undoubtedly carry the sand grain over a longer distance than the boulder. You probably carry around a bunch of sand grains in your shoes every day! Just like carrying the boulder requires more energy for you to lift and carry than a sand grain, the geological processes—often involving the flow of water—that move these sediments also “get tired” and drop the sediments depending on how much energy is required to move the sediment. More energy is required to move sediments like boulders and cobbles than sands and clays. Consequently, large sediments can only be moved by fast moving streams and in places where gravity can act strongly, like in the mountains. To the contrary, even the slowest moving stream can usually move sand and clay particles. Most clay particles only settle out of water when it reaches a standstill, like in deep lakes or the ocean. By understanding this relationship between sediment size and the energy required to move sediments of different sizes, we can reasonably guess the environment based on sediment size.

Confluence of two streams in Denali National Park, Alaska. Note that the stream in the foreground is brown in color because it has a much higher sediment load (likely sand, silt, and clay) than the blue-colored stream in the background. Also notice all of the large pebbles on the shore in the foreground; if these were buried and lithified (i.e., turned to rock), they would form a layer of conglomerate. Photograph by Jonathan R. Hendricks.
Sedimentary features
Sediments are doubly informative about paleoecology because sedimentary features are also commonly preserved and these give additional clues about where and how the sediments were deposited. For example, mudcracks form in an environment where sediments are wet part of the time but also dry out completely at other times, leaving the cracking features we can see in image below. Hummocks and swales are additional features commonly found and form due to waves washed sediments back-and-forth in shallow marine settings. Another common feature, ripples, indicate a nearshore, shallow environment right at the interface of marine and terrestrial ecosystems. Features like these provide additional clues about physical aspects of the environment where they formed.

Sedimentary bedding structures in Devonian-aged siltstones in Central New York State. Note that some beds cut across others, indicating a moderate amount of wave energy and therefore relatively shallow water conditions. The presence of fossils like brachiopods indicates that this place was once a marine habitat.
Fossils as sedimentary particles
From a sedimentological standpoint, fossils themselves can be considered particles just like pieces of sand and clay. As such, the orientation and disarticulation of fossils can provide yet more information about the environment of a fossil assemblage. Many fossils, and parts of fossils, are elongate (one physical dimension is considerably longer than the other two) and this makes them behave predictably in environments where there is flowing water. Examine the image below. What direction do you think water was flowing in this environment?

Approximately ten million year old Turritella shells, many of which are oriented by water flow in the same direction. Fossils are from the Gatun Formation of Panama. Image by Cristina Robins.
In this case, water was flowing approximately left to right, or right to left. We know this because elongate particles tend to orient perpendicular to the direction of flow. Using other information, like changes in sediments or habitat transitions, can help determine which way was shoreward and seaward. Other types of fossil particles, like clam shells, can provide additional information. Clams shells, which we will consider to be shaped like cups, can either have their opening fall upwards or downwards. When examining a fossil assemblage that includes clams, like in the image below, we can look at the proportion of upwards and downward shells to understand how much the assemblage was moved around before it was ultimately deposited and became a part of the fossil record. This can give us clues about how much time-averaging or habitat mixing might have occurred and gives us a sense of how intense waves and storm events were in the environment.

Fossil bivalve shells preserved in the Pliocene Purisima Formation at Capitola Beach near Santa Cruz, California. Note that most of the shells have their concave interiors pointing upwards. Image by Jonathan R. Hendricks.
Disarticulation of fossils
The final way we’ll consider fossils as sedimentary particles is through the process of disarticulation. Disarticulation describes the extent to which the skeletal components of an organism have remained intact as a fossil relative to how the components were oriented in the living organism. Different organisms can exhibit different degrees of disarticulation. For example, a snail really only has one preservable hard part, its whorled shell—the operculum, which the snail uses to cover the opening in its shell, is found occasionally but not very often—whereas a clam has two valves. Thus, it is not common to talk about the articulation of a snail, but when clams are found with both valves as they were when the clam was living, it tells us that the clam likely was buried quickly after it died, before taphonomic processes could separate the valves.

Plio-Pleistocene-aged mollusk fossils from southern Florida. Note that most (but not all) of the bivalve shells are disarticulated. Photograph by Jonathan R. Hendricks.
Many organisms have more than two components. For instance, most vertebrates, like fish, horses, and us, have dozens if not hundreds of bones that can potentially be separated from each other through taphonomic processes. Just like the clam, when we find a vertebrate skeleton that is articulated, meaning it has all of its bones in a life-like orientation, it tells us that the skeleton was rapidly buried and undisturbed after death. To the contrary, a jumble of bones indicates a greater role of taphonomy. What types of processes do you think could lead to the disarticulation of a vertebrate skeleton? In marine settings, wave action can break apart skeletons and so too can other organisms through scavenging. Similarly, on land, scavengers will often move and destroy skeletal remains. In both settings, though somewhat more commonly on land, organismal remains may simply erode away into dust, leaving only partial bones or no fossils at all.
Fossil organisms
In recreating the stories of past environments, we have yet to discuss the lead characters: the fossil organisms themselves. As has been discussed so far, there is much an abundance of information available before we ever consider the fossils but using what we know about organismal biology often helps complete the story. At the start of this section, you made predictions about temperatures based on seeing palm trees or a polar bear. This is a very general use of what you already know about organismal biology. You know that palm trees are found in warm environments and polar bears in cold environments. When paleoecologists consider more of the specific information about organisms, details of the paleoenvironment can also become more specific. Like the palm tree and polar bear example, if you were to find the fossilized remnants of a coral reef you would likely be able to guess that the past environment was a marine setting. After all, applying the assumption of taxonomic uniformitarianism, you probably know that today coral reefs are found in marine ecosystems. Other than its marine origin, what might a coral be able to tell you about the past environment? To answer this question, we’ll consider two aspects of the coral, it’s morphological characteristics and its chemical composition.

A fossilized coral reef from the 125,000 year old Cockburn Town Member on San Salvador Island, Bahamas. Photo by Jansen Smith.
Morphological characteristics
Morphology describes the form or shape of a living thing and can be thought of as a collection of pliable characteristics molded by ecological and evolutionary processes so that an organism can survive and reproduce in a specific habitat (learn more about evolutionary processes in the chapter on Evolution and the Fossil Record). This means that the morphologies of corals we find in the fossil record can help us interpret the environment in which the coral was living, again by applying the concept of taxonomic uniformitarianism. In particular, the shape of a coral colony is highly informative about the exposure of the coral to wave energy—you can think of this as the frequency and size of waves pushing against a coral—and we can observe living corals to understand this relationship. Looking at the 3D models of the corals below, which do you think are best suited for an environment with high wave energy? And low wave energy? What features did you consider when distinguishing between them?
Fossil specimen of the scleractinian coral Septastrea crassa from the Pliocene Tamiami Fm. (Pinecrest Beds) of Sarasota, Florida (PRI 45562). Specimen is from the research collections of the Paleontological Research Institution, Ithaca, New York. Length of specimen is approximately 23.5 cm.
Fossil scleractinian “pancake” coral Siderastrea pliocenica from the Pliocene Tamiami Formation of Lee County, Florida (PRI 76859). Specimen is from the research collections of the Paleontological Research Institution, Ithaca, New York. Longest dimension of specimen is approximately 14 cm.
Modern specimen of the scleractinian coral Montastraea cavernosa from off Monroe County, Florida (PRI 13355). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Maximum dimension of specimen is approximately 11 cm.
Generally, mounded (e.g., Montastrea cavernosa; see 3D model above) and flattened (e.g., Siderastrea pliocenica; see 3D model above) corals are found in areas with high wave energy and more delicate, branched forms (e.g., Septastrea crassa; see 3D model above) occur in areas with lower wave energy. This pattern is largely driven by the damage that waves can cause to corals. With the more delicate forms, waves can more easily break off branches and this can kill the coral, expose it to diseases, or, at the least, require energy—which may have otherwise been used by the coral to grow bigger or to reproduce—to be repaired. On the contrary, the species with flattened forms and, even more so, with mound forms are resistant to wave-induced breakage. As is typically the case in ecology, if an organism is well suited for a particular condition (like mounded corals being able to cope with wave impacts), it usually comes at the cost of something else. This is referred to as a trade-off. In this case, mounded corals tend to grow more slowly than their branched counterparts. Consequently, when mounded and branched forms coexist (in a low-wave environment), branched forms can often crowd out the mounded forms because they grow more quickly. Those branches can block out the sunlight required for corals to grow, which gives them a competitive advantage. Because these differences exist and can be validated by examining living corals, a paleontologist studying corals can pinpoint the location on a reef where the coral was living. The paleontologist will have a good sense of how much wave energy was in the system and can reasonably determine if the habitat was a lagoon, a barrier reef, or another coral habitat.

Corals growing in the Palmyra Atoll National Wildlife Refuge in the Pacific Ocean. Image by Jim Maragos for the U.S. Fish and Wildlife Service (public domain).
Much like tree rings and ice cores, corals grow incrementally and their hard calcium-carbonate skeleton can be studied to understand environmental factors like temperature, salinity, and even the number of days during a year. Corals grow a tiny amount each day and the amount they grow can be linked to changes in the tidal cycle and seasons. By examining these growth bands in corals from the past, it is possible to count the number of days in the year. Studying a rugose coral from upstate New York, paleontologist John W. Wells was able to determine that, during the Silurian and Devonian, days on earth were approximately two-hours shorter than they are today and there were about 400 days in a year! That is, in the same total amount of time it takes us to travel once around the sun, the earth used to rotate so much faster that there were 35 more days in a year, though each day would have been a little shorter. It was only possible to draw this conclusion by first understanding the ecology of how corals grow today and then applying that understanding to the fossilized corals.

Heliophyllum halli from the Middle Devonian Moscow Formation of Erie County, New York (PRI 70755). This is one of the genera of rugose corals studied by Wells, though is not one of his specimens (the location(s) of Wells' specimens are currently unknown). Insets show details of fine growth lines (left; likely not daily growth lines, however) and possible annual banding (right). This specimen is from the collections of the Paleontological Research Institution, Ithaca, New York.
Chemical Composition
The high-resolution record in coral skeletons also makes them ideal candidates for biogeochemical analysis. As we will explore in the next session, biogeochemistry is the study of the integrated biological, geological, and chemical processes (e.g., rock, water, and carbon cycles) and reactions that dictate the dynamics of natural environments. Much like we discussed earlier with ice cores and tree rings, corals and other organisms with hard parts can be studied at a chemical level to gain even more information about the environment. Because corals can be sampled on a yearly basis (see the image below), they can be studied to understand short term changes in climate and water chemistry. Continue reading about biogeochemistry in the next section to learn about how carbon, oxygen, and other isotopes from organisms like corals can be used to improve our understanding of life in the past.

Cores taken from living corals showing annual growth bands. "(a) X-ray positive print of top 50-cm section of Porites coral core slice from Tantabiddi Reef, Western Australia (AIMS ID = TNT03BS2), collected in October 2008, showing unusual density band in 1998; (b) X-ray positive print of 65-cm section of Porites coral core slice from Thevenard Island, Western Australia (AIMS ID = THV01BS3), collected in October 2008, showing growth disruption in 1958; and (c) digitally enhanced photograph of top 46-cm section of Porites coral core slice from Pandora Reef, GBR (AIMS ID = PAN37AS2) collected in August 2012. Image illustrates annual luminescent lines of varying intensity." Figure and caption from Lough and Cantin (2014).
Concept check: See what you know!
What is a proxy?
In paleoecology, a proxy an indirect means to assess another variable of interest. Proxies are used because it is often easier to measure a correlated, indirect variable than the actual variable of interest.
What are three examples of proxies?
Tree rings, ice cores, species occurrences, morphological features, biogeochemical data
What is the relationship between sediment particle size and energy in a depositional environment?
Larger sediments (e.g., boulders, cobbles, gravels) require more energy to move and so are found in environments like alluvial fans and rivers. Smaller sediments are more easily moved and so are found in lower energy environments like lakes, lagoons, and the deep ocean.
If in a fossil coral assemblage you find only individuals with intricate, branching morphologies, what inferences could you make about the environment where the corals lived?
Branching corals are prone to breaking in high wave energy and so are typically found in lower energy environments like lagoons and back reef zones.
How does knowing the morphology of an organism help interpret the environment where it lived?
Organisms tend to be adapted to the environments where they live, meaning their morphologies reflect environmental conditions and differences in morphology can be used to describe differences in environment.
References
Bottjer, D. J., M. L. Droser, and D. Jablonski. 1988. Palaeoenvironmental trends in the history of trace fossils. Nature, 333: 252-255.
Chappell, J. 1980. Coral morphology, diversity and reef growth. Nature, 286: 249-252.
Lough, J. M., and N. E. Cantin. 2014. Perspectives on massive coral growth rates in a changing ocean. The Biological Bulletin, 226: 187-202.
Martinelli, N. 2004. Climate from dendrochronology: latest developments and results. Global and Planetary Change, 40: 129-139.
Mills, P. C. 1983. Genesis and diagnostic value of soft-sediment deformation structures—a review. Sedimentary Geology, 35: 83-104.
Petit, J. R., J. Jouzel, D. Raynaud, N. I. Barkov, J. M. Barnola, I. Basile, M. Bender, J. Chappellaz, M. Davis, G. Delaygue, and M. Delmotte. 1999. Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica. Nature, 399: 429-436.
Wells, J. W. 1963. Coral growth and geochronometry. Nature, 197: 948-950.
Wells, J. W. 1966. Paleontological evidence of the rate of the Earth’s rotation. Pp. 70-81, in: The Earth-Moon System, B. G. Marsden & A. G. W. Cameron (eds.), Springer, Boston.
Usage
Unless otherwise indicated, the written and visual content on this page is licensed under a Creative Commons Attribution-NonCommercial-Share Alike 4.0 International License. This page was written by Jansen A. Smith. See captions of individual images for attributions. See original source material for licenses associated with video and/or 3D model content.
