


Chapter contents:
Paleoecology
– 1. Paleoenvironmental Reconstruction
– 2. Biogeochemical Analysis and Paleoecology
– 3. Predation in Paleoecology
–– 3.1 Insect Herbivory in the Fossil Record ←
–– 3.2 Drilling Predation in the Fossil Record
–– 3.3 Dinosaur Predation in the Fossil Record
Above image by E. Bradford Walker; Creative Commons Attribution 3.0 United States license.
{kind=link}
Overview
Predation, the consumption of one organism by another, takes many forms and can include plants being eaten by insects. That is, herbivory can be considered a special case of predation where the prey is a plant. Though insects themselves do not have a very complete fossil record—even if there are some excellently preserved fossil insects in amber (i.e., hardened tree resin)—trace fossils give us an indication of how some insects behaved. Most commonly, insect trace fossils take the form of indications of predation on plants and their leaves. By studying the presence and frequency of these interactions, it is possible to estimate the diversity of past insects and their behaviors. Once again, this area of study draws heavily on comparisons to modern plant-insect interactions to ground truth the evidence we see in the fossil record.

A small fly preserved in amber. Inset shows the same specimen under ultraviolet light. Image by Zacharie Grossen; Creative Commons Attribution-Share Alike 4.0 International license.
{kind=link}
Types of insect damage
Leaf damage from insects can occur in a variety of forms, ranging from galls to chew marks on the leaf margin. When studying living organisms, these traces can be directly linked to the insects leaving the trace through observation and experimentation. Once the trace is connected to a particular type of insect, the presence of the insect can be surmised from the presence of the trace alone. For example, if you have ever worked in a garden, you might inspect the leaves of different plants to determine whether any pest insects are damaging them. Since garden pests are relatively well studied, a quick Google search can help you identify the insect making the traces on your garden plants (and how to get rid of them). Although the emphasis is different in paleoecology, a similar process is used to identify different types of insects in the fossil record.

A caterpillar (left) and beetle (right) eating the margins of leaves. Left photograph by Toby Hudson; Creative Commons Attribution-Share Alike 3.0 Unported license. Right photograph by Cfp: Creative Commons Attribution-Share Alike 2.5 Generic, 2.0 Generic and 1.0 Generic license.
{kind=link}
{kind=link}
Before turning to examples from the fossil record, we will first examine a few different types of insect herbivory on modern plants. An understanding of interactions between modern plants and insects can help contextualize and interpret observations from the fossil record by utilizing the assumption of taxonomic uniformitarianism.
Chewing traces
The first traces we will consider come from the action of an insect chewing on a leaf or stem. When an insect chews on a plant, usually on the leaf, it can leave holes or notches on the leaf. These traces are typically left by insects like grasshoppers, caterpillars, or beetles. Some insects will chew up so much of the leaf that the only remnants are the leaf’s tough veins; this is called defoliation or skeletonization. Other insects use their chewing mouthparts to bore or tunnel into a plant’s leaf or stem. These actions tend not to destroy the entire leaf, such as in defoliation, and instead leave characteristic tunnels in the plant material. Although we have only considered these types of traces in a general sense, keep in mind that different insects make slightly different traces that can be distinguished from each other. For example, one caterpillar might leave holes in the center of a leaf and another might chew away parts of the leaf edge. These subtle differences help identify the type of insect leaving the trace.

A leaf with holes made by chewing insects. Photograph by Forest and Kim Starr; Creative Commons Attribution 3.0 United States license.
.jpg){kind=link}
Galls
Rather than directly consuming plant material, some insects insert their larvae into stems or leaves and the larvae then consume the plant. This type of behavior commonly results in the formation of a gall in the plant. The gall is a reaction by the plant—in some ways similar to swelling in the human body—to isolate and reduce the effect of the intrusive insect. In the United States, one of the most common examples of galls is in the genus Solidago, or goldenrod. In goldenrod, the presence of a larvae, often of a moth or butterfly species, induces the growth of a large gall. These large galls also commonly attract predators, including wasps and woodpeckers. Whereas woodpeckers will peck open the gall and eat the larval insect, the wasps tend to lay their own eggs in the gall. When the wasp eggs hatch, the wasp larvae then consume the larvae of the moth or butterfly. In many ways, galls can be a microhabitat of their own and while this level of resolution is not always present in the fossil record, the observation of these modern interactions hints at the types of ecological interactions that may have occurred in the past.

A living goldenrod plant with a gall (left) and a split open gall showing the fly larvae responsible for the gall (right). Left photograph by R.A. Nonenmacher; Creative Commons Attribution Share-Alike 4.0 International license. Right photograph by Beatriz Moisset; Creative Commons Attribution Share-Alike 2.5 Generic license.
{kind=link}
{kind=link}
Sucking traces
The last type of leaf damage we will consider is from the behavior of insects sucking on plant tissues to consume their nutrients. This sucking behavior, commonly conducted by aphids, scale insects, and leaf hoppers, can result in spotting or stippling patterns on leaves and even the formation of misshapen leaves and fruits. In modern plants, there may also be discoloration corresponding to the damage, though these color changes are less likely to be preserved in the fossil record.

Damage from scale insects on tree leaves. Notice the yellow discoloration where the insects were sucking on the plant tissues to gain nutrients. Photograph by Doug Beckers; Creative Commons Attribution Share-Alike 2.0 Generic license.
.jpg){kind=link}
Trace examples in the fossil record
When examining the traces of insect herbivory on plants in the fossil record, the assumption of taxonomic uniformitarianism is critical. Insects, aside from cases of exceptional preservation, have a very poor fossil record. Consequently, it would be very challenging to identify the particular insect that created a particular herbivory trace without information from modern observations. Even modern observations have limited applicability, as the further back in time we go, the less similar species are to those living today. Reflecting this challenge, traces are often treated as unique entities themselves—this is also true of other kinds of traces, like drill holes, burrows, and tracks. In this treatment, individual types of traces receive their own name, similar to the way a species gets a name. These names can be useful because they provide a systematic way to talk about the traces; however, we will consider the traces in a more descriptive way, as this is fundamentally what forms the basis of the assigned names. Examine the traces in the image below. Referring to the section on “Types of insect damage,” can you identify whether the traces were from sucking, galls, or chewing?

Insect damage on a fossilized leaf from the Cretaceous of Kansas. The nearly round holes at the left and top of the leaf were likely created by a hole-feeding insect. Photograph by Wilson44691; Wikimedia Commons (public domain).
{kind=link}
Plant-insect interactions across the Cretaceous-Paleogene extinction
Mass extinctions are times of wide, sweeping change in life on Earth, as numerous species die out and new species arise to take their places during the thousands of years that follow. One pattern that has emerged from the study of Earth’s major mass extinctions is that specialist species are more likely to go extinct than their generalist counterparts. Specialists, which are organisms that are very well adapted to do a small number of things, are reliant on a particular set of environmental conditions, like the existence of a certain host species, a specific temperature range, or a predictable amount of rainfall. When those conditions change abruptly, like during a mass extinction, specialist species tend to die out. To the contrary, generalists, which are more flexible in the conditions they can tolerate, are more likely to survive because of their flexibility. For a closer look at this pattern, we can turn to plant-insect interactions across the Cretaceous-Paleogene mass extinction that occurred approximately 66 million years ago—the same event that led to the extinction of non-avian dinosaurs.

"A spectrum of plant–insect associations from the Williston Basin of southwestern North Dakota. Associations range from the earliest Paleocene at upper left, 14.4 m above the KT boundary, and continue to the older associations of the latest Cretaceous at lower right, 85.5 m below the boundary. All material is from the Denver Museum of Nature and Science (DMNH) or the Yale Peabody Museum (YPM). Following each plant host are, respectively, morphotype number (indicated by the prefixes HC or FU) (17), specimen number, DMNH locality number (loc.), and meter distances from the KT boundary as in Fig. 2 (17). Damage types are indicated by the prefix DT (Fig. 2). (Scale bars: solid 1 cm, backslashed 0.1 cm.) (a) Two linear mines with oviposition sites (arrows), following secondary and then primary venation, terminating in a large pupation chamber (DT59), on the dicot Paranymphaea crassifolia (FU1),DMNH20055, loc. 563,14.4 m. (b) Single gall (DT33) on primary vein of Cercidiphyllum genetrix (Cercidiphyllaceae, FU5), DMNH 20042, loc. 562, 8.4 m. (c) Free feeding (DT26) on Platanus raynoldsi (Platanaceae, FU16), DMNH 20035, loc. 2217, 1.3 m. (d) Skeletonization (DT61) on a probable lauralean leaf (HC32), DMNH 19984, loc. 2097, 31.4 m. (e) Multiple galls (DT33) on Trochodendroides nebrascensis (Cercidiphyllaceae, HC103),DMNH19976, loc. 1489,33.7 m. (f) Initial phase of a serpentine mine (DT45) on Marmarthia pearsonii (Lauraceae, HC162), DMNH 7228, loc. 2087, 36.9 m. (g) Cuspate margin feeding (DT12, arrow) on Metasequoia sp. (Cupressaceae, HC35), DMNH 13108, loc. 567, 56.8 m. (h) Serpentine leaf mine (DT43) assigned to the Nepticulidae (Lepidoptera), on unidentified Rosaceae (HC80), YPM 6367a, loc. 567, 56.8 m. (i) Hole feeding pattern (DT57) on an unknown genus of Urticales (HC81), DMNH 19539, loc. 2203, 56.8 m. (j) General skeletonization (DT16) on Erlingdorfia montana (Platanaceae, HC57), DMNH 11013, loc. 571, 61.7 m. (k) Large scale-insect impressions (DT53) centered on primary veins of E. montana, DMNH 18829b. loc. 571, 61.7 m. (l) Slot hole feeding (DT8) on an unidentified genus of Platanaceae (HC109), DMNH 18658, loc. 434, 88.5 m." Figure and caption from Labandeira et al. (2002) in the Proceedings of the National Academy of Sciences under the PNAS License to Publish (Copyright (2002) National Academy of Sciences).
Working in southwestern North Dakota, Conrad Labandeira, Kirk Johnson, and Peter Wilf examined more than 13,000 fossil plant specimens across the Cretaceous-Paleogene boundary for evidence of damage from insects. The plant specimens they studied, which were preserved in lake and stream deposits, included representatives from all major groups of plants from the time and are from layers of rock representing 2.2 million years spanning the extinction boundary.
The researchers found only five insect body fossils but 51 unique types of plant-insect interactions. To be counted as an herbivory trace, four criteria had to be met: (1) presence of reaction tissue produced by the plant; (2) details of the trace margins linked to anatomy of the insect mouth parts; (3) a stereotypical pattern, position, or size and shape of the damage; and, (4) specificity in the interaction between an insect and plant. If a trace met these criteria, the researchers then categorized the type of trace (e.g., hole or margin feeding, skeletonization, mining, galling) and the specific morphology of the trace. Each of the different 51 traces was also determined to be generalist, intermediate, or specialist based on whether the trace was found in many, few, or one plant host.
Based on what you know so far, do you expect more generalist, intermediate, or specialist interactions to be present both before and after the extinction?
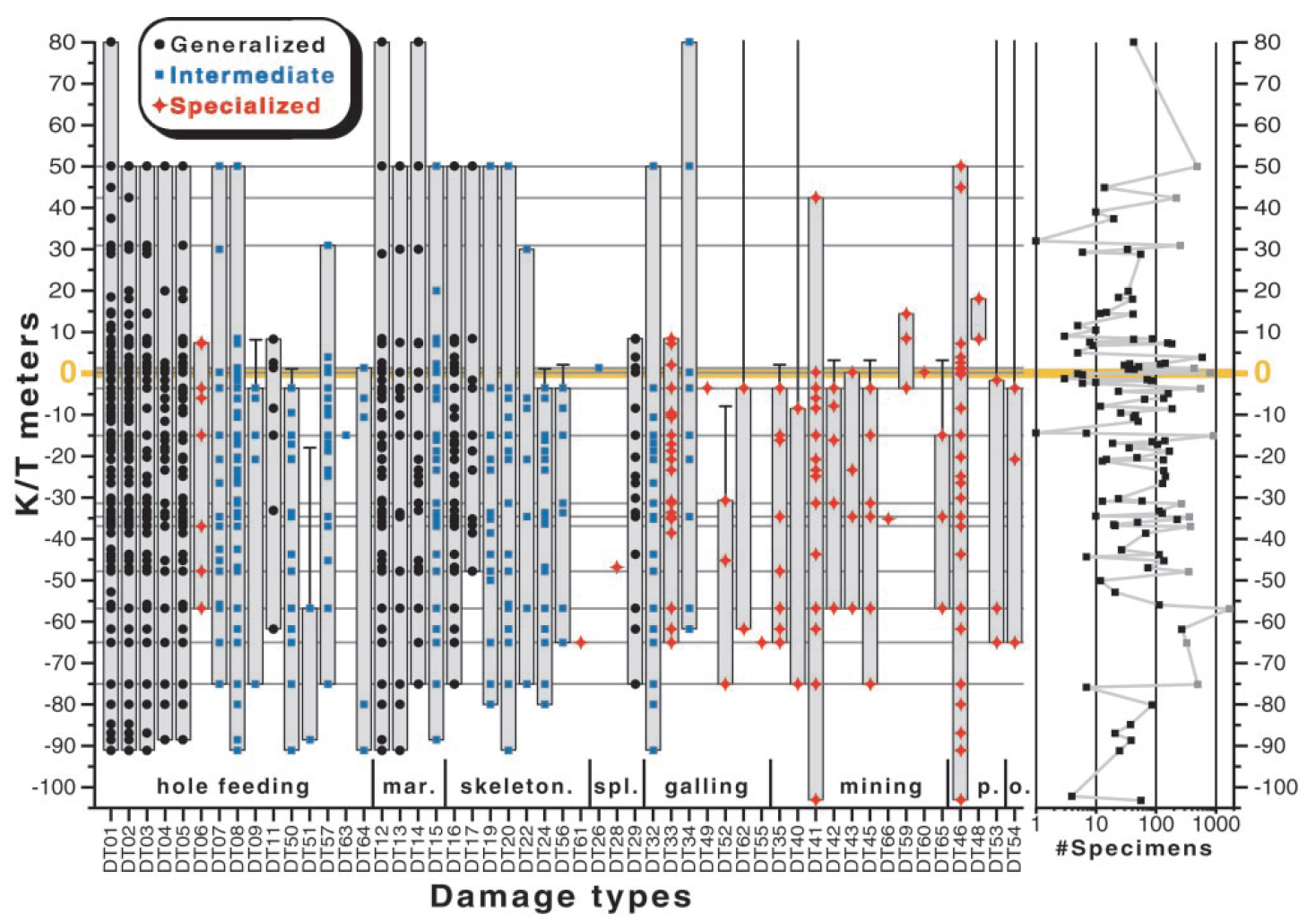
"Stratigraphic and sampling data for the 51 insect-mediated damage types from a 183-m composite section straddling the KT boundary (orange bar), from the Williston Basin of southwestern North Dakota (Table 1 and Table 2, which is published as supporting information on the PNAS web site). The estimated time duration for this interval is 2.2 million years (my) before present, of which 0.8 my are assigned to the postboundary interval and 1.4 my are allocated to the preboundary interval (2). The Cretaceous strata are within the Hell Creek and Fort Union formations, and the Paleocene strata are entirely within the Fort Union Formation (2, 17). Placement of the zero datum is at the KT boundary, not the Hell Creek/Fort Union formational contact, because the contact isdiachronous with respect to the KT boundary timeline (17). Depicted are the raw presenceabsence data for damage types, categorized by functional feeding group and subgroup at the bottom, and representing 106 discrete horizons. Eight damage types have single stratigraphic occurrences (dots); the ranges of those with multiple stratigraphic occurrences are shown as vertical shaded bars, categorized by host specificity from the color scheme at upper left (see text). Confidence intervals of 50% are provided for the 14 damage types whose regional last appearances occur before the boundary, using a method that accommodates nonrandom distributions of fossil horizons and sampling intensity by allowing recovery potential to vary stratigraphically (39) (see text). Of the 14 specialized damage types with multiple stratigraphic occurrences that disappear at or below the KT boundary, six (42.9%) reappear during the latest Paleocene to middle Eocene of Wyoming and Utah (27, 30). All associations are extant today. Four confidence intervals that exceed our sampling range are shown without end bars. Total specimen frequencies are given at right, including all plant organs; note logarithmic scale. The gray horizontal lines in the main figure and corresponding gray squares at right indicate the 14 horizons with more than 200 specimens of identified dicot leaves, used in Fig. 3; meter levels for these floras are: 75.0, 65.0, 56.8, 47.8, 36.9, 34.6, 31.4, 15.0, 3.6, 0.2, 1.3, 30.9, 42.4, 50.0. Abbreviations: mar., margin feeding; skel., skeletonization; spl., specialized external feeding types; p., piercing and sucking; o., oviposition. Limitations on graphical presentation cause the flora at 0.2 m, which is Paleocene, to appear within the orange line representing the KT boundary." Figure and caption from Labandeira et al. (2002) in the Proceedings of the National Academy of Science under the PNAS License to Publish (Copyright (2002) National Academy of Sciences).
As you may have guessed, generalist interactions were much more likely to be found on both sides of the extinction boundary. In fact, all twelve of the generalist traces found in the pre-extinction Cretaceous were also found in the post-extinction Paleogene. This was not the case for intermediate and specialist species. Ten of the sixteen (62.5%) intermediate traces crossed the boundary. Only six of twenty (30%) specialist traces do so. [Three traces were not included because of uncertainty in the data.] Considering the types of traces, most of the generalists were hole (n=6) or margin (n=3) feeders that simply consumed a portion of leaf tissue. Intermediate species were also often hole feeders (n=8), but a large number were also prone to skeletonizing (n=5) leaf tissue. Two intermediate species did induce galls in their plant hosts; however, galling was much more commonly a trace associated with specialists (n=5). The most common forms of specialist interactions were, however, characterized as mining traces (n=10). From an ecological standpoint, this breakdown in trace type and specialization is linked to the manner in which the insect utilizes its plant counterpart. Recall the section above on types of insect damage: what might explain the connection between trace types and specialization?
As described above, galling and mining traces tend to be associated with insect reproduction and, to an extent, shelter. With these traces, insects often embed themselves (or their offspring) within the plant’s leaf or stem. As a result of environmental conditions and the specific needs of the insects, strong associations between plants and insects are often formed, just as in the Solidago gall example above. When considering hole and margin feeding, the associations between insects and plants is often much weaker. Though some species will form specific associations—for example, caterpillars of monarch butterflies are commonly found on milkweed—many hole and margin feeding insects are capable of consuming the tissues of many different types of plants, hence the generalist tendency. Grounded in comparisons to modern plant-insect interactions, these remarkable traces of 66-million-year-old interactions reveal the broad pattern that generalist species are more likely to survive times of extinction than their specialist counterparts. Thinking of today, as we undergo the sixth mass extinction in Earth’s history, we can expect a similar pattern to play out.
Concept check: See what you know!
List three types of damage made to plant stems and leaves by insects.
Types of damage include, but are not limited to, chewing traces, galls, and sucking traces.
How do galls differ from other types of plant damage?
Chewing and sucking traces are made, primarily, by insects directly consuming nutrients from the plant. Galls are typically formed by a plant in response to a disturbance or intrusion. Insect induced galls are often made after an insect lays eggs inside the plant (and the hatched larvae will eat from the plant).
In terms of diet breadth, what is the difference between a specialist and a generalist?
A specialist diet typically includes only one, or a few, food sources, which the specialist is efficient at consuming. A generalist diet includes many more sources than a specialist and generalists tend to be less efficient at consuming any of those sources compared to a species specializing on the same food source.
During an extinction, is a specialist or a generalist more likely to survive?
A generalist is more likely to survive an extinction because it has more flexibility in how it secures resources.
Which type of feeding trace is more likely to be associated with a generalist herbivore insect: margin feeding or galling?
Margin feeding is more likely to be associated with a generalist insect herbivore. The behavior of eating leaf margins does not require the specificity that is often required in an interaction that produces galls.
References and Further Reading
Labandeira, C. C., K. R. Johnson, and P. Wilf. 2002. Impact of the terminal Cretaceous event on plant–insect associations. Proceedings of the National Academy of Sciences, 99: 2061-2066.
Labandeira, C. C., K. R. Johnson, and P. Lang. 2002. Preliminary assessment of insect herbivory across the Cretaceous-Tertiary boundary: major extinction and minimum rebound. Geological Society of America Special Paper, 361: 297-327.
Labandeira, C. C., and E. D. Currano. 2013. The fossil record of plant-insect dynamics. Annual Review of Earth and Planetary Sciences, 41: 287-311.
Weis, A. E., and W. G. Abrahamson. 1986. Evolution of host-plant manipulation by gall makers: ecological and genetic factors in the Solidago-Eurosta system. The American Naturalist, 127: 681-695.