

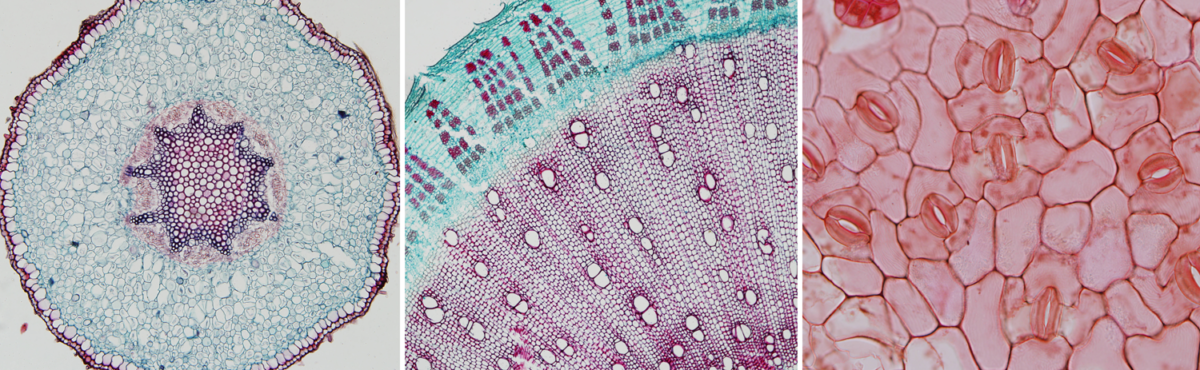
Feature image. Plant anatomy sections. Left: Cross section of a polyarch root of Aeglopsis chevalieri, a plant in the citrus family (Rutaceae) of angiosperms. Center: Cross section of a woody stem of kenaf (Hibisicus cannabinus, an angiosperm). Right: Epidermis of a leaf of redgal (Morinda royoc, an angiosperm). Credits: Aeglopsis chevalieri Swingle (SV422), Hibiscus cannabinus L. (C8), and Morinda roioc (F24) (University of Miami Swingle Plant Anatomy Reference Collection, not under copyright in the United States). Images modified from originals.
What are vascular plants (tracheophytes)?
Most land plants are tracheophytes, or vascular plants. Their defining feature is the presence of vascular tissue, or specialized tissue that conducts food (phloem) and water (xylem). When considered in a phylogenetic context or in the fossil record, tracheophytes can be more specifically defined by the origin of a type of water-conducting cell known as a tracheid. The first evidence for tracheids shows up in the fossil record in the Silurian period. Tracheophytes later diversified structurally, with many key innovations appearing by the end of the Devonian period, such as leaves and wood. The dominant life stage in most vascular plants is the sporophyte, or the diploid, spore-producing plant.
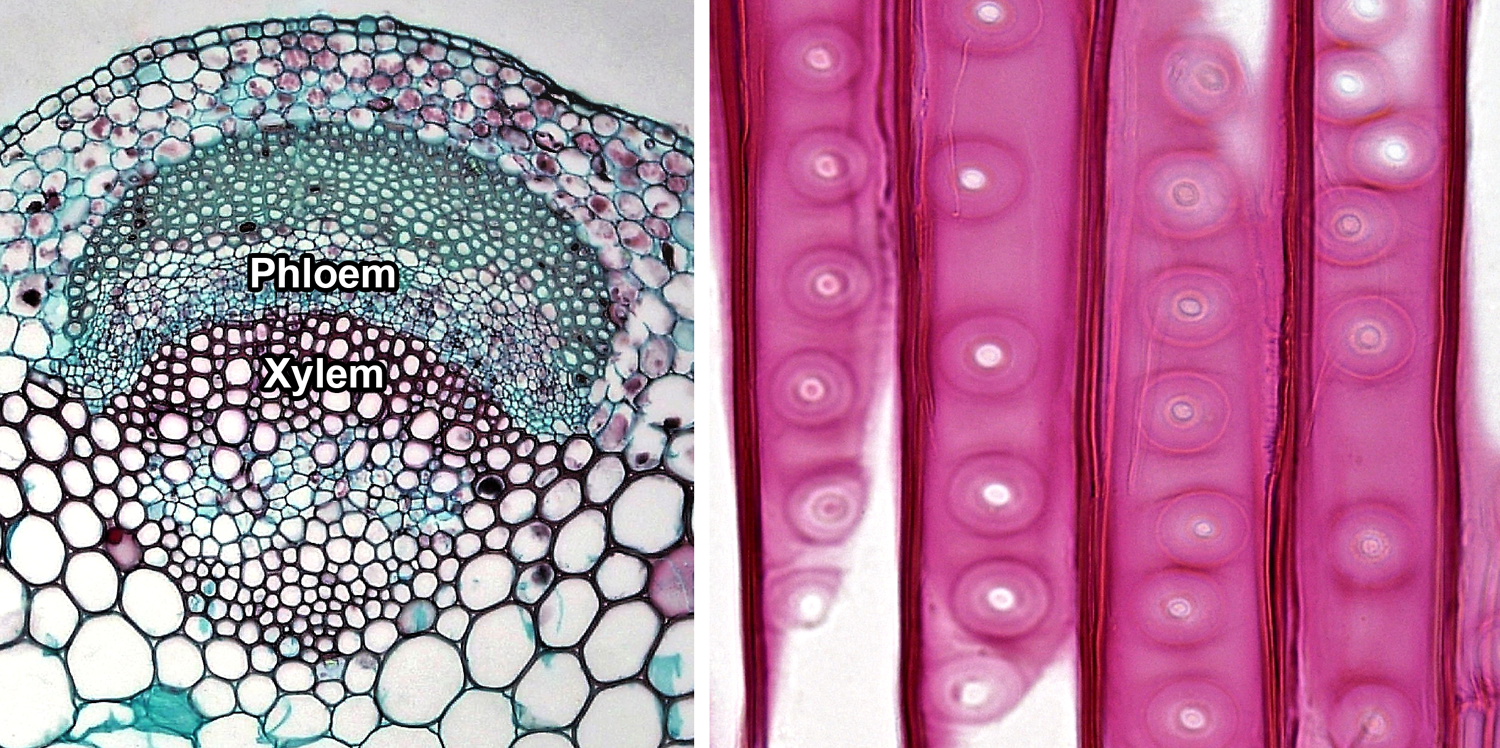
Vascular tissue. Left: Cross section of a vascular bundle in the stem of clover (Trifolium, an angiosperm) showing food-conducting phloem and water-conducting xylem. Right: Detail of portion of the tracheids in pine (Pinus, a conifer) wood, radial section (longitudinal section on the radius of the axis); note the characteristic circular-bordered pits typical of conifer tracheids. Credits: Cortex and pith in Trifolium stem and Circular bordered pits in Pinus wood (Fayette A. Reynolds M.S./Berkshire Community College Bioscience Library, via flickr, CC0 1.0/Public Domain Dedication).
Living tracheophytes belong to one of two major sister groups: 1) The lycophytes (clubmosses and relatives); and 2) The euphyllophytes (ferns, horsetails, and seed plants). The lycophytes, ferns, and horsetails are free-sporing (seedless) vascular plants sometimes collectively called pteridophytes. These plants share certain plesiomorophic or ancestral structural and developmental characteristics. Thus, it is sometimes useful when discussing the comparative sporophyte structure of vascular plants to contrast the paraphyletic pteridophytes with the monophyletic spermatophytes, or seed plants. Furthermore, while angiosperms (flowering plants) are a group of seed plants, they have many of their own unique structural characteristics that set them apart from gymnosperms (the group made up of non-angiospermous seed plants).
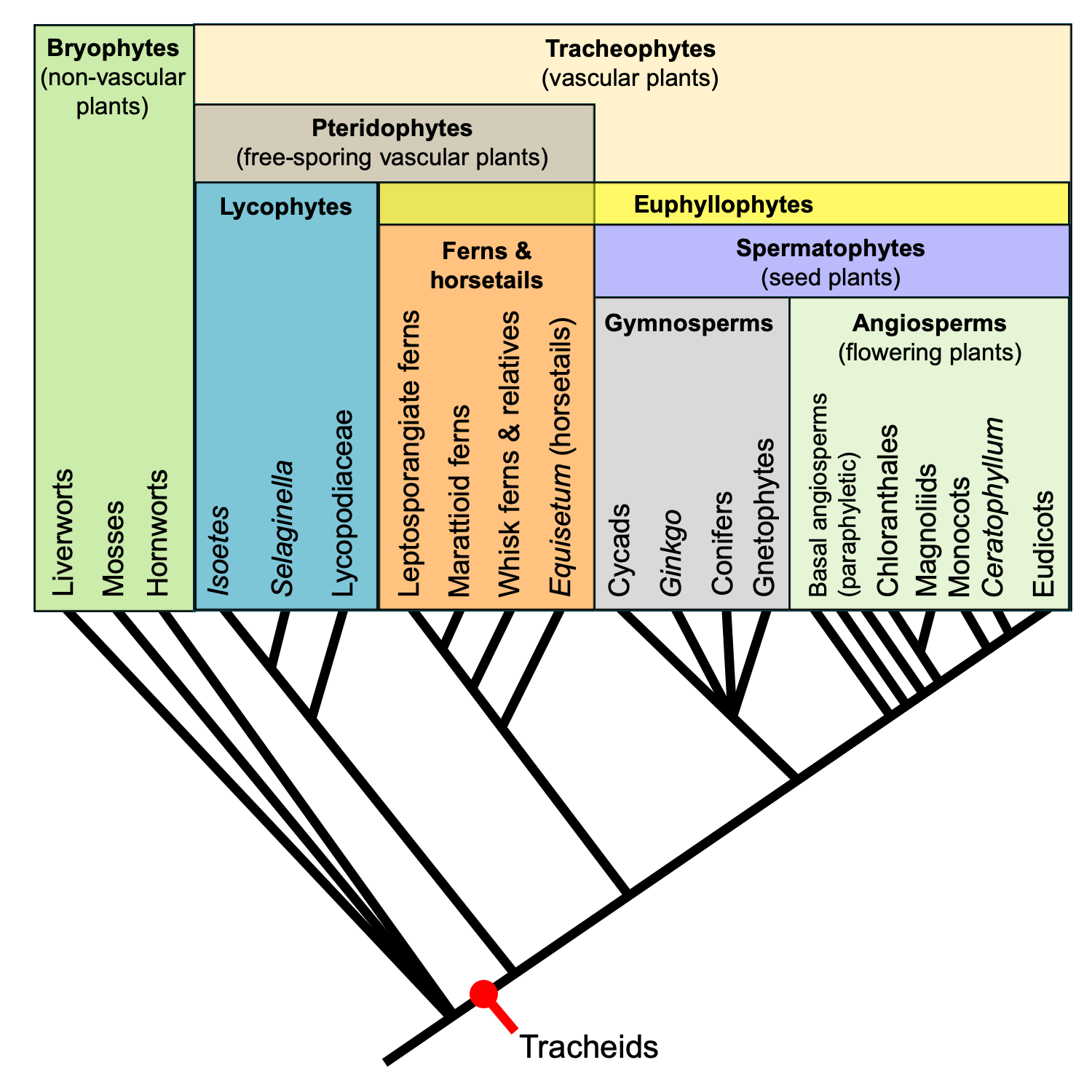
Relationships among major extant land plant groups. This phylogenetic tree shows the relative relationships among extant land plants, with emphasis on vascular plant groups. Note that the relationships among extant gymnosperm groups is shown as unresolved. Recent molecular phylogenetic analyses indicate that extant gymnosperms are monophyletic; this contradicts the traditional view that extant gymnosperms are a paraphyletic group. Regardless, gymnosperms are paraphyletic when extinct groups are included. Credit: E.J. Hermsen (DEAL).
Overview of sporophyte structure
The study of plant structure includes morphology, the study of form, and anatomy, the study of internal structure (cells, tissues, etc.). Paleobotany uses both of these disciplines in order to analyze fossil plants.
Organs & organ systems of the plant body
The vascular plant sporophyte (hereafter, simply called the plant) includes three basic categories of non-reproductive, or vegetative, organs. These are the roots, stems, and leaves. Roots are organs that typically occur belowground and anchor the plant in the soil. They also function in absorbing water and sometimes in storage. Stems are often (although not always) aboveground structures. Stems bear the leaves, and also serve as conduits between roots and leaves. Finally, leaves are lateral appendages on the stem. Their main function is photosynthesis. A stem and its leaves together are known as a shoot.
Plant organs can be grouped into two organ systems. All the shoots (stems and leaves) of a plant are together known as the shoot system, whereas all the roots together comprise the root system. Sporangia (spore-producing structures) are borne on modified leaves or shoots; thus, sporangium-bearing organs can also be considered part of the shoot system.
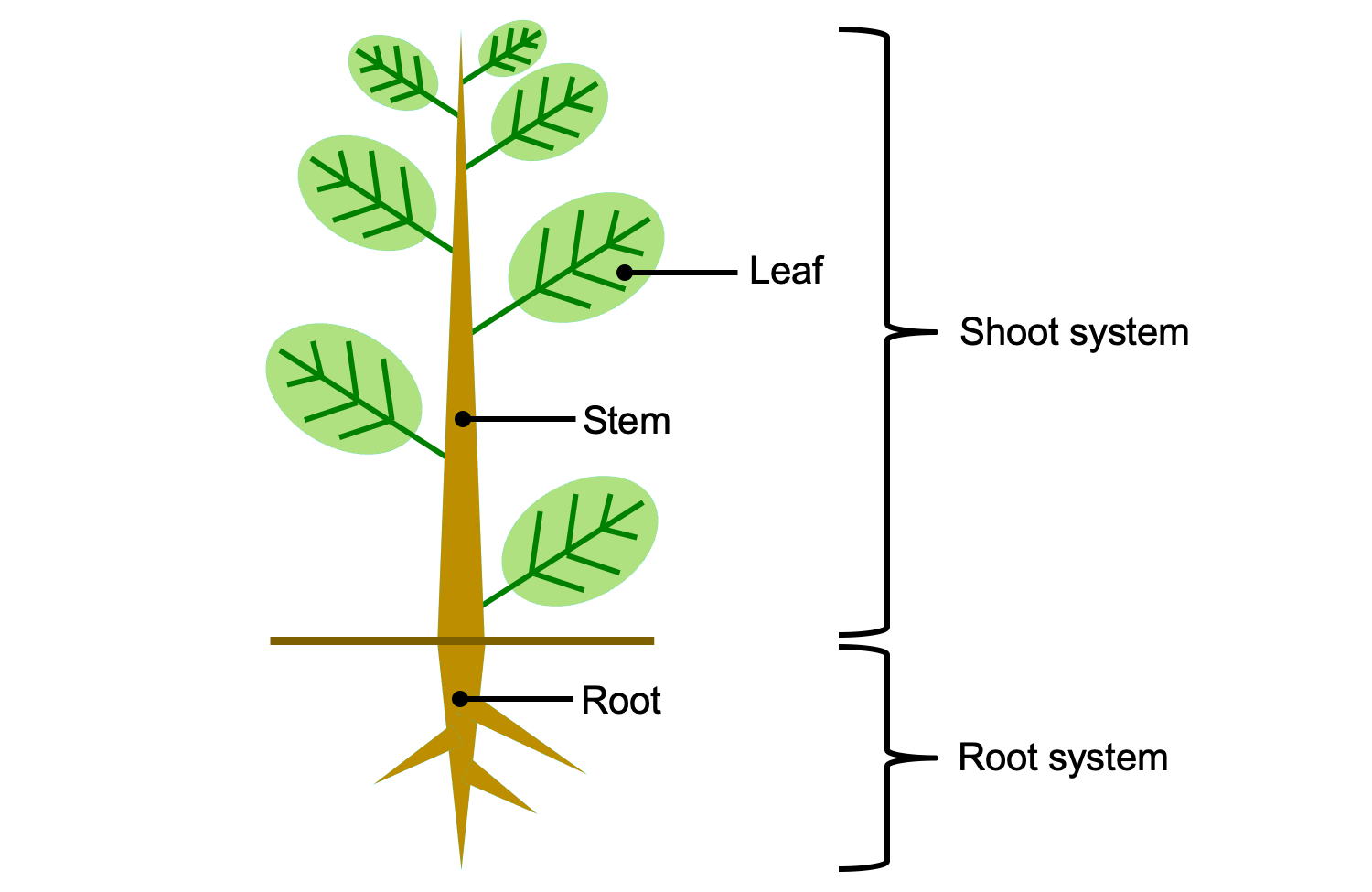
Vegetative organs and organ systems of the vascular plant sporophyte. The shoot system includes al of the stems, leaves, and reproductive structures (sporangia, cones, flowers, etc.). The root system includes all of the roots. Credit: E.J. Hermsen (DEAL).
Growth
Growth is the process by which a plant increases in size (length, height, or girth) through cell division (adding new cells) and cell expansion. Active cell division is typically concentrated in regions known as meristems. As cells are distanced from the meristems by new growth, they begin to differentiate. Differentiation is the process by which a cell takes on a specific identity and function in the plant body. Once a cell is fully differentiated, or mature, it often does not—and in many cases cannot—divide further.
Vascular plant sporophytes have two major categories of meristems that contribute to growth: apical and lateral. However, other types of meristems may contribute to growth of stems and expansion of leaves (e.g., intercalary meristems, marginal meristems).
Apical meristems
Apical meristems are meristems at the tips of roots and stems that add length to these organs; growth from apical meristems is known as primary growth. Primary growth also produces the leaves. The primary meristems—protoderm, ground meristem, and procambium—include partially differentiated cells. The primary meristems can be found in regions still undergoing growth between the apical meristem proper and the fully mature tissues of the stem, leaf, and root. They correspond to the three tissue systems of vascular plants (discussed below).
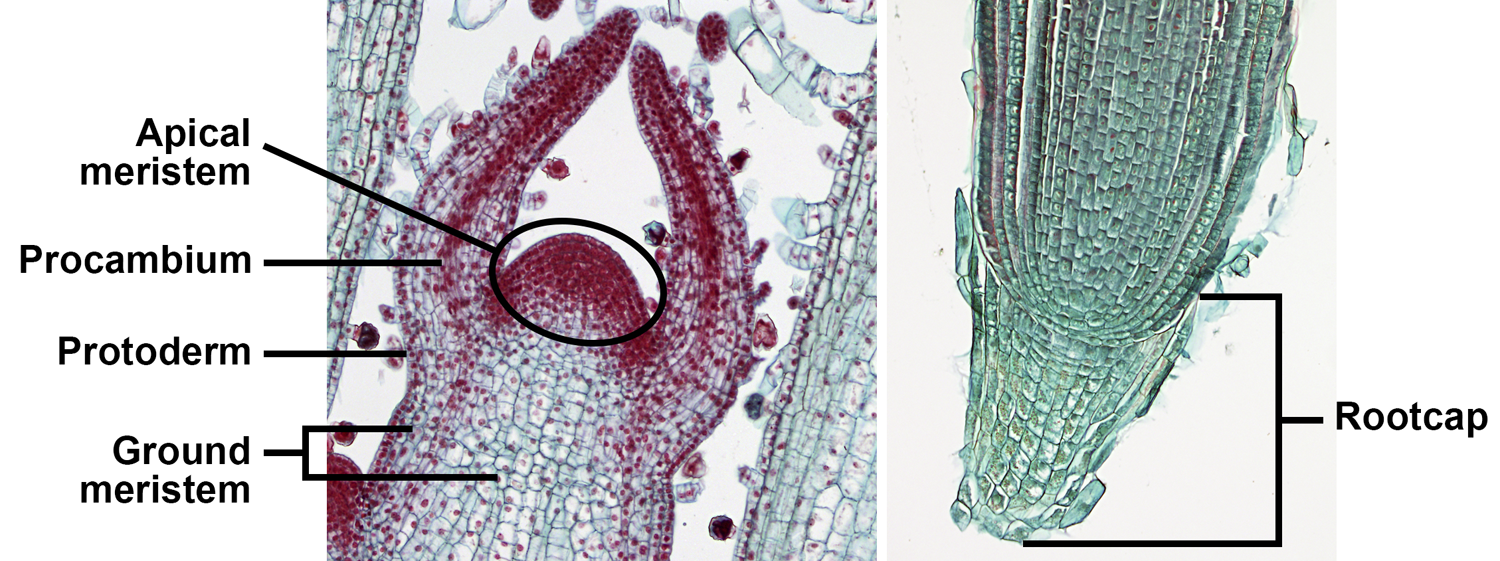
Apical and primary meristems. Left: Longitudinal section of the shoot apex of coleus (Plectranthus, an angiosperm), showing the shoot apical meristem and primary meristems; the horn-like projects to either side of the apical meristem are developing leaves. Right: Longitudinal section of the root apex of wheat (Triticum); the root apical meristem is immediately above the root cap. Credits: Coleus shoot tip and wheat root tip (BlueRidgeKitties, via flickr, CC BY-NC-SA 2.0). Images modified from originals.
Lateral meristems (cambia)
Lateral meristems, also called cambia or secondary meristems, are located within roots and stems and increase the diameter of these structures. Growth from cambia is known as secondary growth. Living pteridophytes do not have secondary growth. Seed plants may or may not have secondary growth, depending on the taxon.
A woody stem or root has two major concentric regions; wood is surrounded by bark. In a typical woody seed plant, two types of lateral meristems contribute cells to these regions. The vascular cambium is a layer of dividing cells between the wood and the bark. It produces vascular tissue to its interior (xylem) and to its exterior (phloem), thus contributing to the growth of both the wood and the bark. The cork cambium develops within the bark and most notably adds layers of protective cork to the bark.
Bark encompasses the entire region outside of the vascular cambium. In addition to tissues produced by the cambia, it may also include tissues produced by primary growth. Wood is made up entirely of xylem (more specifically, secondary xylem, or xylem produced by secondary growth).
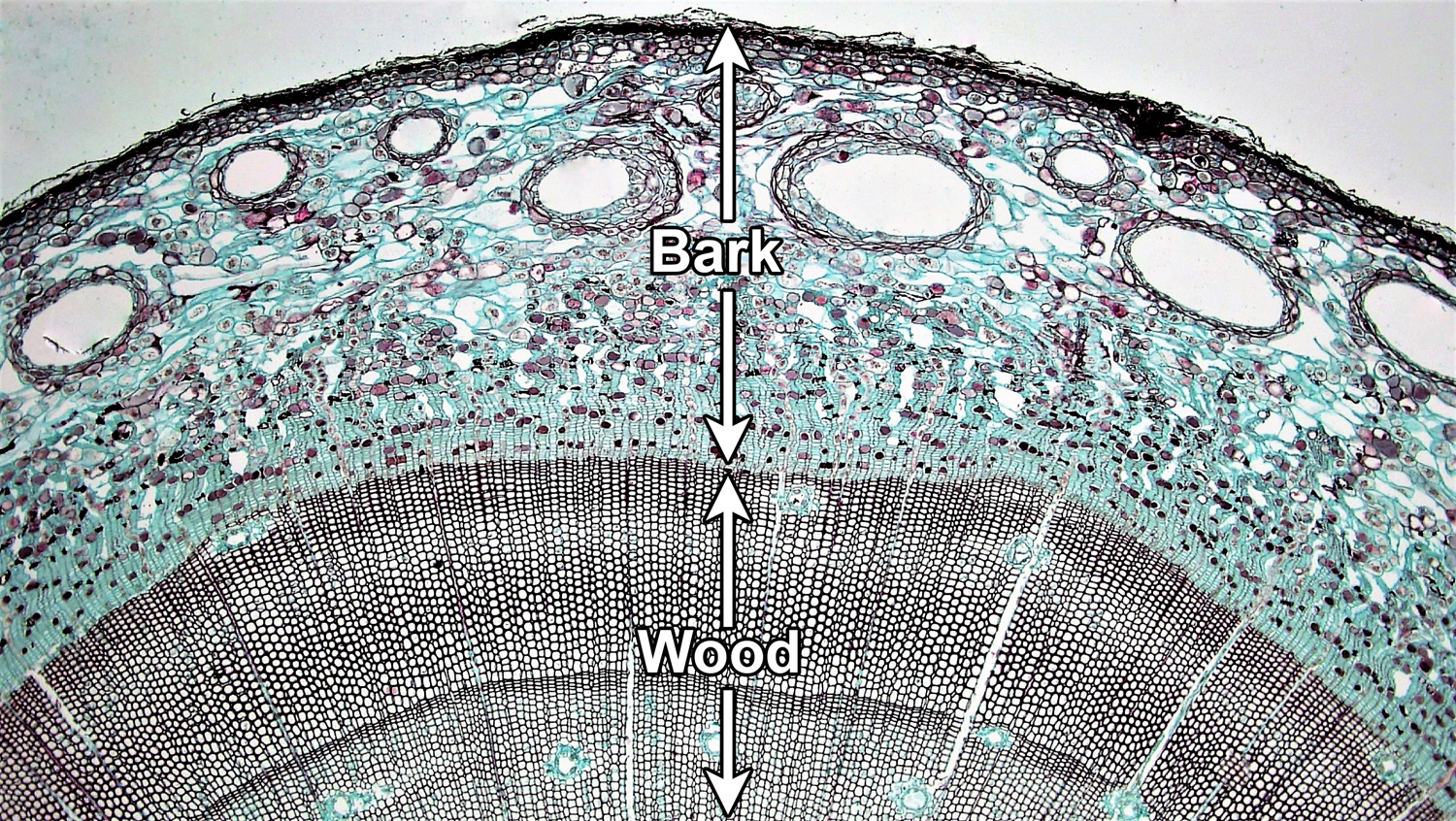
Vascular cambium. Cross section of a woody pine (Pinus) stem. The vascular cambium is a layer of dividing cells located between the wood and the bark. Note that the vascular cambium produces cells both to its inside and its outside; the cells are produced in neat rows. In this young stem, much primary tissue is still present in the bark and there is no well-developed cork layer. For a good example of cork, see the cross section of birthwort (Aristolochia) periderm below. Credit: Four year Pinus (Fayette A. Reynolds M.S./Berkshire Community College Bioscience Image Library, via flickr, CC0 1.0/Public Domain Dedication). Image modified from original.
Tissues & tissue systems
Tissues are regions of cells in the plant body that are functionally and physically connected to one another. There are two basic categories of tissues in the plant body, simple and complex. Simple tissues are made up of one type of cell, whereas complex tissues include more than one type of cell. Tissues can also be classified as primary tissues or secondary tissues. Primary tissues are produced by growth from the apical and primary meristems, whereas secondary tissues are produced by growth from the cambia.
Tissues can further be grouped into tissue systems. Tissue systems may include more than one type of tissue. The three tissue systems and their tissues are:
Dermal tissue system
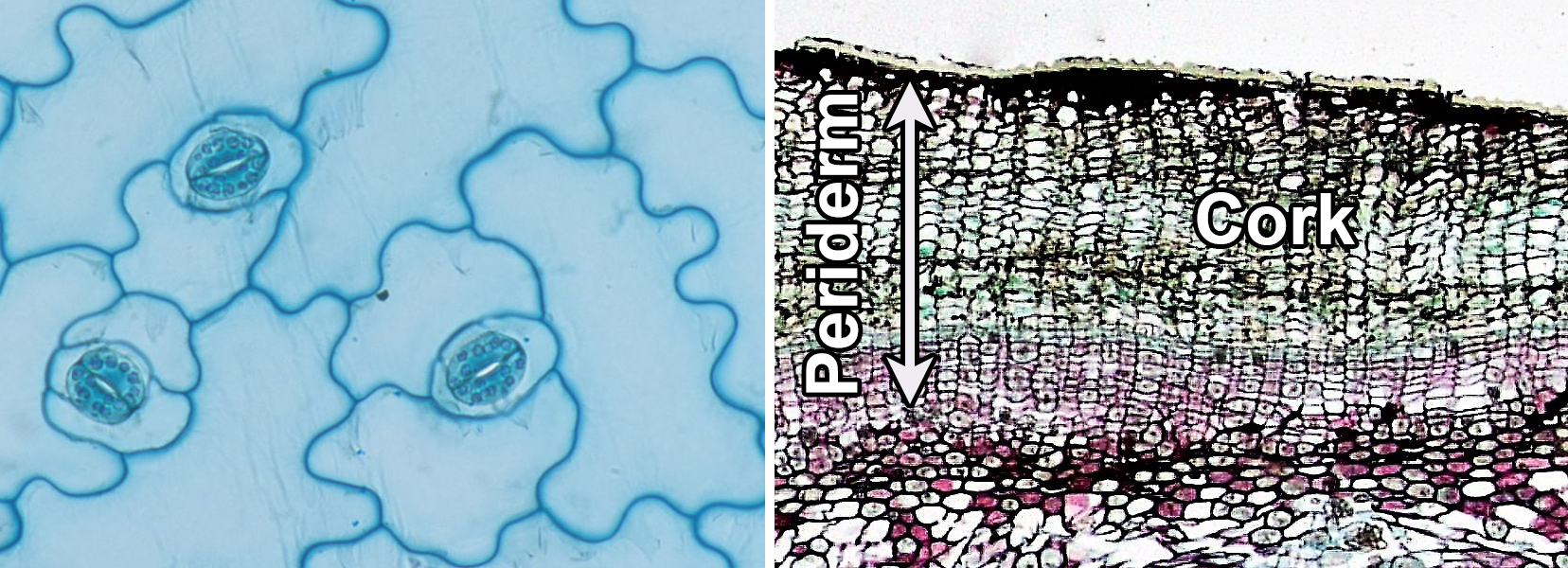
The dermal tissue system covers the outer surface of the plant body. The primary dermal tissue is the epidermis, which matures from the protoderm. The epidermis is typically one cell layer thick and primarily made up of epidermal cells. The secondary dermal tissue is the periderm, which is produced by the cork cambium and makes up part of the bark in woody plants. Periderm consists of multiple layers of cells, including outer layers of protective cork cells (which are dead when mature), the cork cambium, and a typically thin inner zone of living cells. The dermal tissues are complex (made up of more than one cell type).
Vascular tissue system
The vascular tissue system (less often called the fascicular tissue system) conducts food and water through the plant body. The two types of vascular tissue are phloem, or the food-conducting tissue, and xylem, or the water-conducting tissue. Phloem and xylem are complex tissues and include conducting cells and other cell types. The conducting cells are sieve elements in the phloem and tracheary elements in the xylem (a tracheid is a type of tracheary element). Vascular tissue can be primary or secondary, depending on its origin. If it matures from the procambium, it is primary; if it develops from the vascular cambium, it is secondary.
In leaves, vascular tissue is often arranged into one or more veins or vascular bundles (strands) that may form complex networks. The veins are surrounded by a region of ground tissue (discussed below) called the mesophyll.
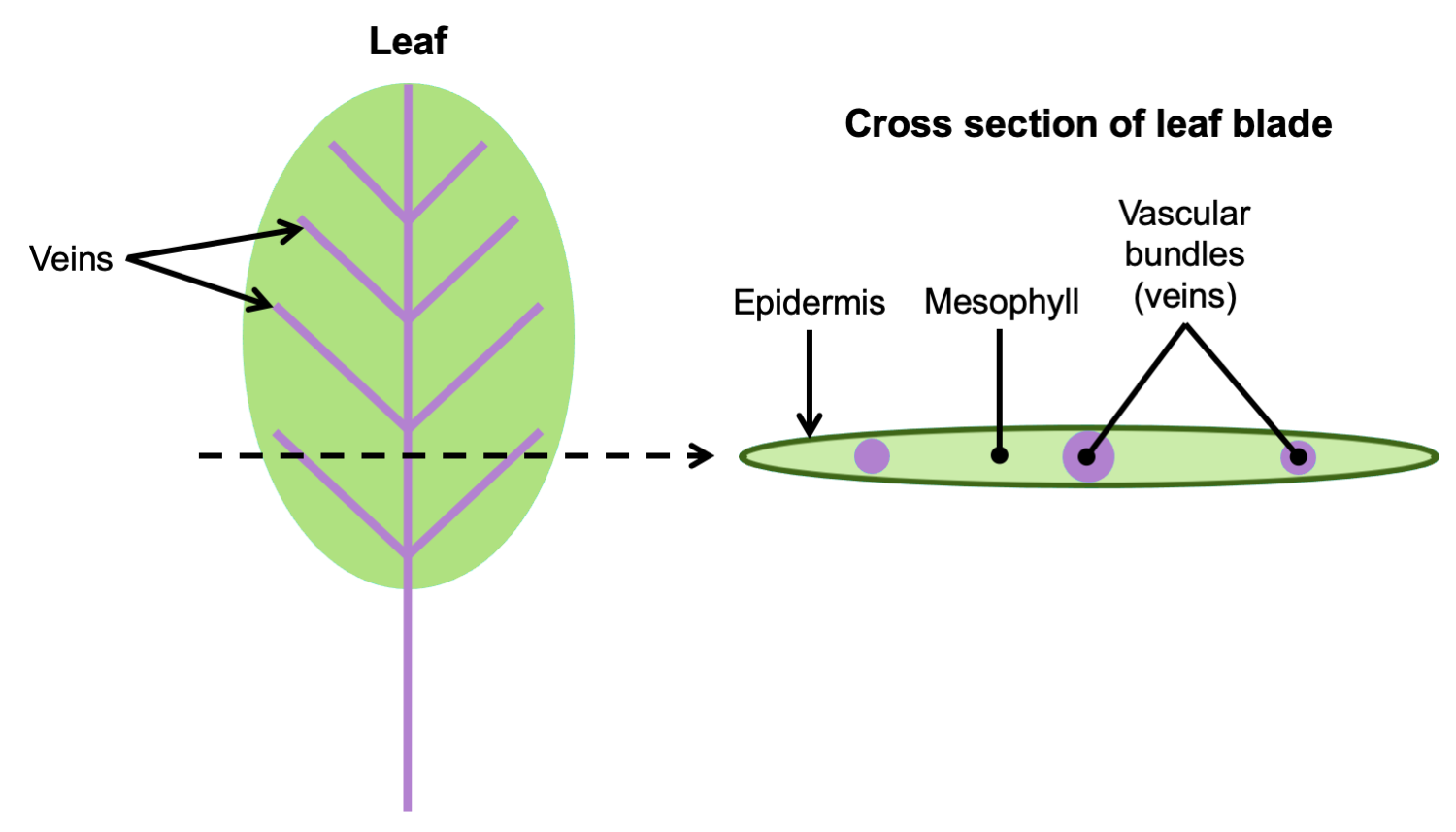
Arrangement of vascular tissue in a leaf. Left: Diagram of a leaf, with the petiole (stalk) and veins in purple; the entire expanded portion of the leaf in which the veins are embedded is the blade. The dotted line indicates where the leaf would be cut to make a cross-section. Right: Cross section of leaf blade showing the vascular bundles embedded in the mesophyll. Credit: E.J. Hermsen (DEAL).
In the stem, the arrangement of primary vascular tissue varies. It can be arranged, for example, in a solid cylinder (protostele), a hollow cylinder surrounding a central pith (siphonostele), or in separate vascular bundles around a pith (eustele). The primary vascular tissue of the root typically takes the form of a protostele (solid cylinder) with a central strand of lobed xylem and phloem between the lobes of xylem.
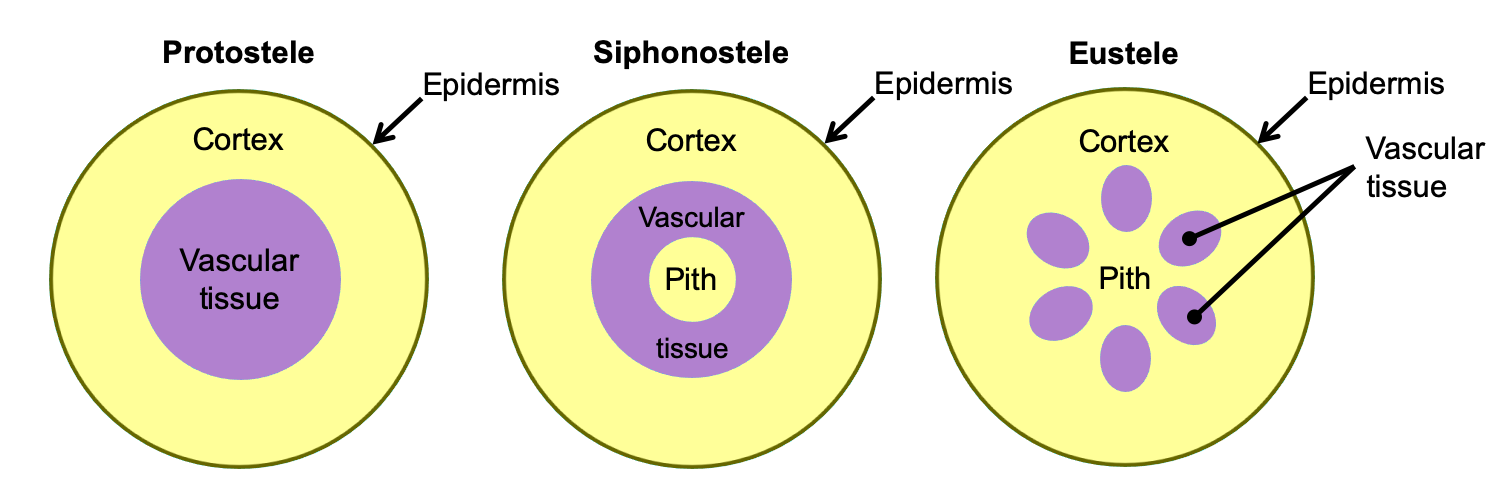
Arrangement of primary vascular tissue in axes (stems and roots). The diagrams above show axes (roots or stems) in cross section (transverse section). Left: Protostele, solid vascular tissue with no pith; this arrangement is typical of roots and of lycophyte stems. Center: Siphonostele, vascular tissue in a ring surrounding a pith; this arrangement is typical of fern stems. Right: Eustele, vascular tissue in bundles surrounding a pith; this arrangement is typical of seed plant stems. Credit: E.J. Hermsen (DEAL).
In stems and roots with secondary growth, the secondary xylem typically forms a continuous ring internal to a continuous ring of secondary phloem. The xylem and phloem are separated by a thin vascular cambium, or layer of undifferentiated dividing cells. Secondary xylem is also known as wood. All tissues external to the vascular cambium are part of the bark. Thus, secondary phloem is part of the bark.
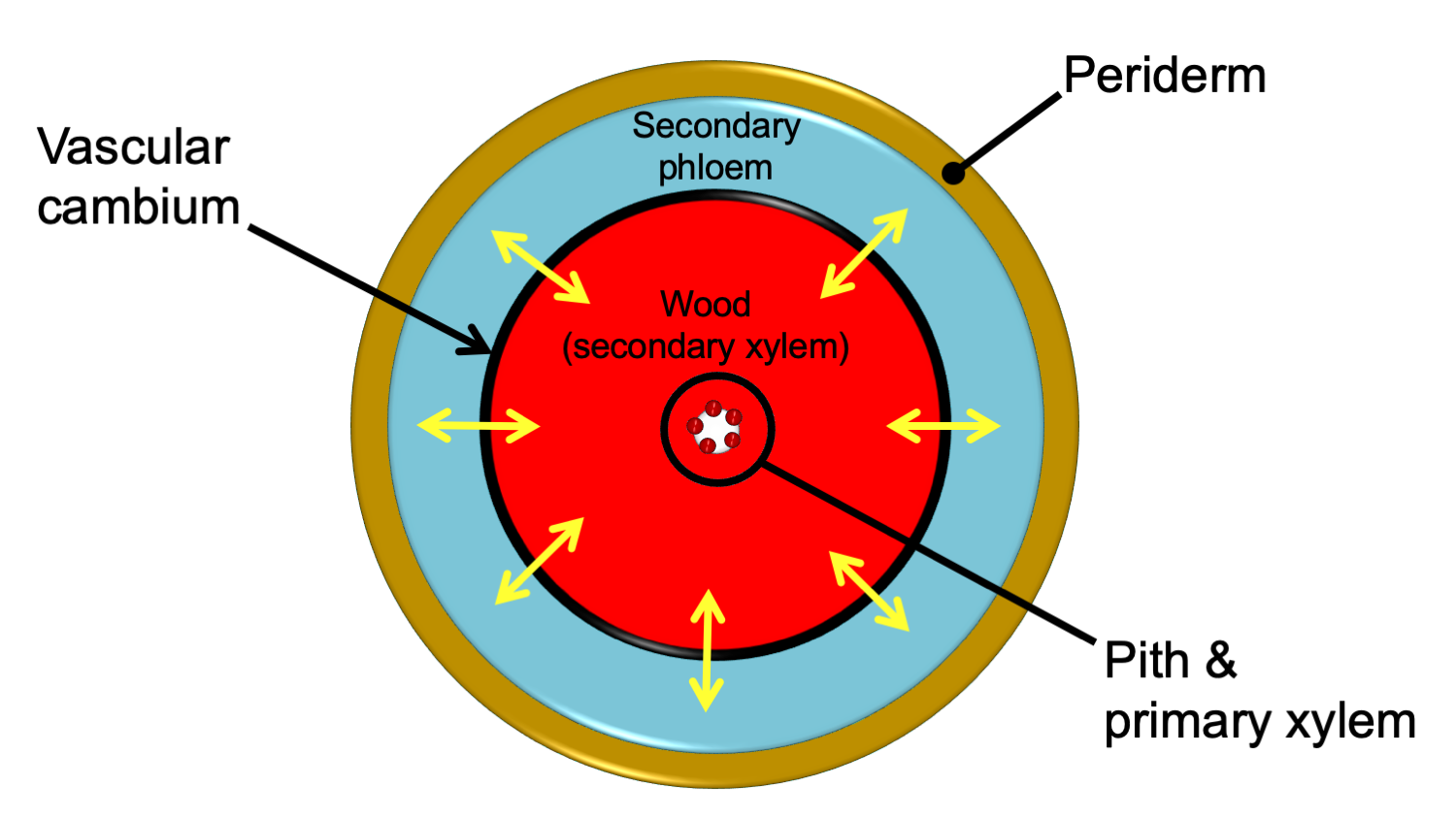
Relative arrangement of secondary tissues in a mature woody stem. This diagram shows the relative arrangement of secondary tissues in a mature seed plant stem in cross section. Note that some of the primary tissues are retained in the center of the woody stem. The yellow arrows show the directions of cell division that add cells to the secondary phloem and secondary xylem. A woody root would be similar in anatomy, except that the center of the stem would be made up of a lobed strand of primary xylem with no pith. In young woody stems, some primary tissues may be retained between the secondary phloem and the periderm (see the cross section of a young pine stem above on this page for an example). These are often crushed and destroyed/sloughed off as the stem ages. Credit: E.J. Hermsen (DEAL).
Ground tissue system
The ground tissue system (also called the fundamental tissue system) fills in the spaces in the plant body between the dermal and vascular tissues; it develops from the ground meristem. Ground tissue is typically primary tissue. It can be found primarily in the following regions of the plant body:
- Cortex: The region between the epidermis and the vascular tissue in the stem and root.
- Pith: The region in the center of the stem, internal to the vascular tissue (not always present).
- Mesophyll: Occurs between and around the vascular bundles of the leaf.
In addition, ground tissue occurs between the vascular bundles in stems with separate vascular bundles, such as stems with eusteles.
There are three types of ground tissue: parenchyma, collenchyma, and sclerenchyma. They are simple tissues. The cells that make up these tissues share the name of their respective tissue type: parenchyma cells, collenchyma cells, and sclerenchyma cells. Parenchyma cells often have thin cell walls, whereas collenchyma and sclerenchyma cells have thicker cell walls. Collenchyma adds support to non-woody stems and leaves, whereas rigid sclerenchyma cells often function in support or protection. Parenchyma is versatile, and functions in photosynthesis, storage, healing, etc.
Note: Parenchyma cells and sclerenchyma cells do not occur only in the ground tissue system. They can also be found in the vascular tissue.
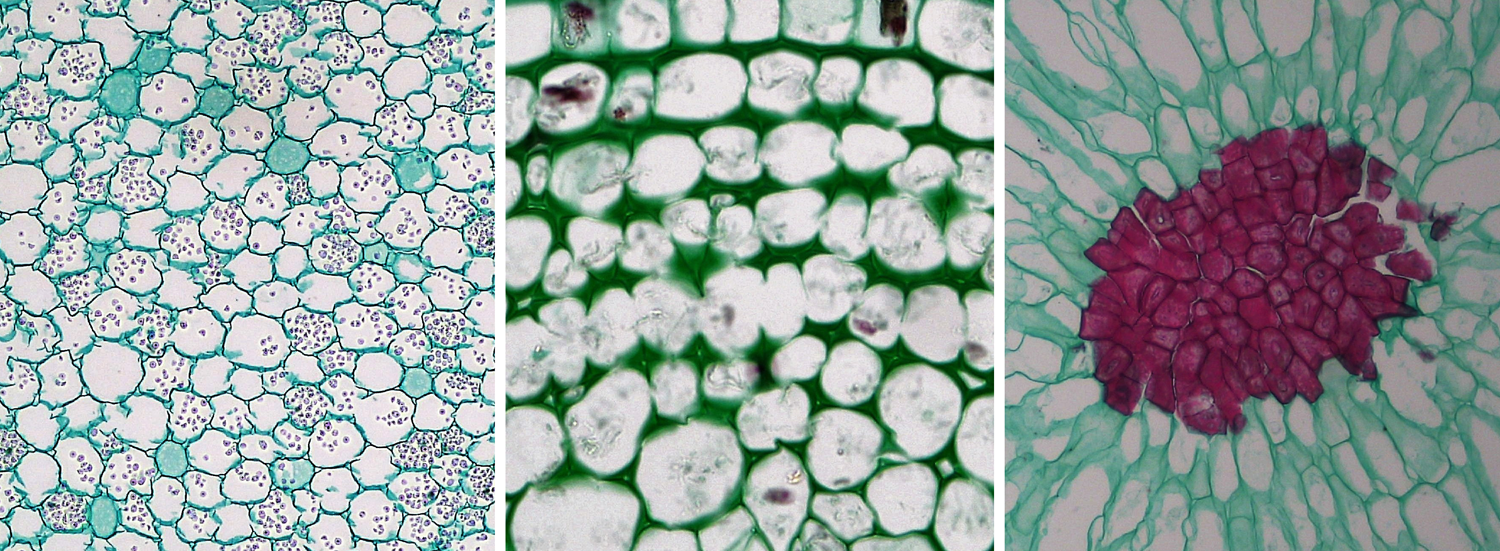
Parenchyma, collenchyma, and sclerenchyma cells. Left: Parenchyma cells from a buttercup (Ranunculus) root, cross section. Center: Collenchyma cells from a sunflower (Helianthus) stem; note the thickened walls of the collenchyma cells, cross section. Right: Sclerenchyma cells from the flesh of a pear (Pyrus); the sclerenchyma cells are a group of stone cells (sclereids) with thick walls that are stained red. Credits: Parenchyma in Ranunculus root, collenchyma in older Helianthus stem, and sclereids in Pyrus (Fayette A. Reynolds M.S./Berkshire Community College Bioscience Image Library, via flickr, CC0 1.0/Public Domain Dedication). Images modified from originals.
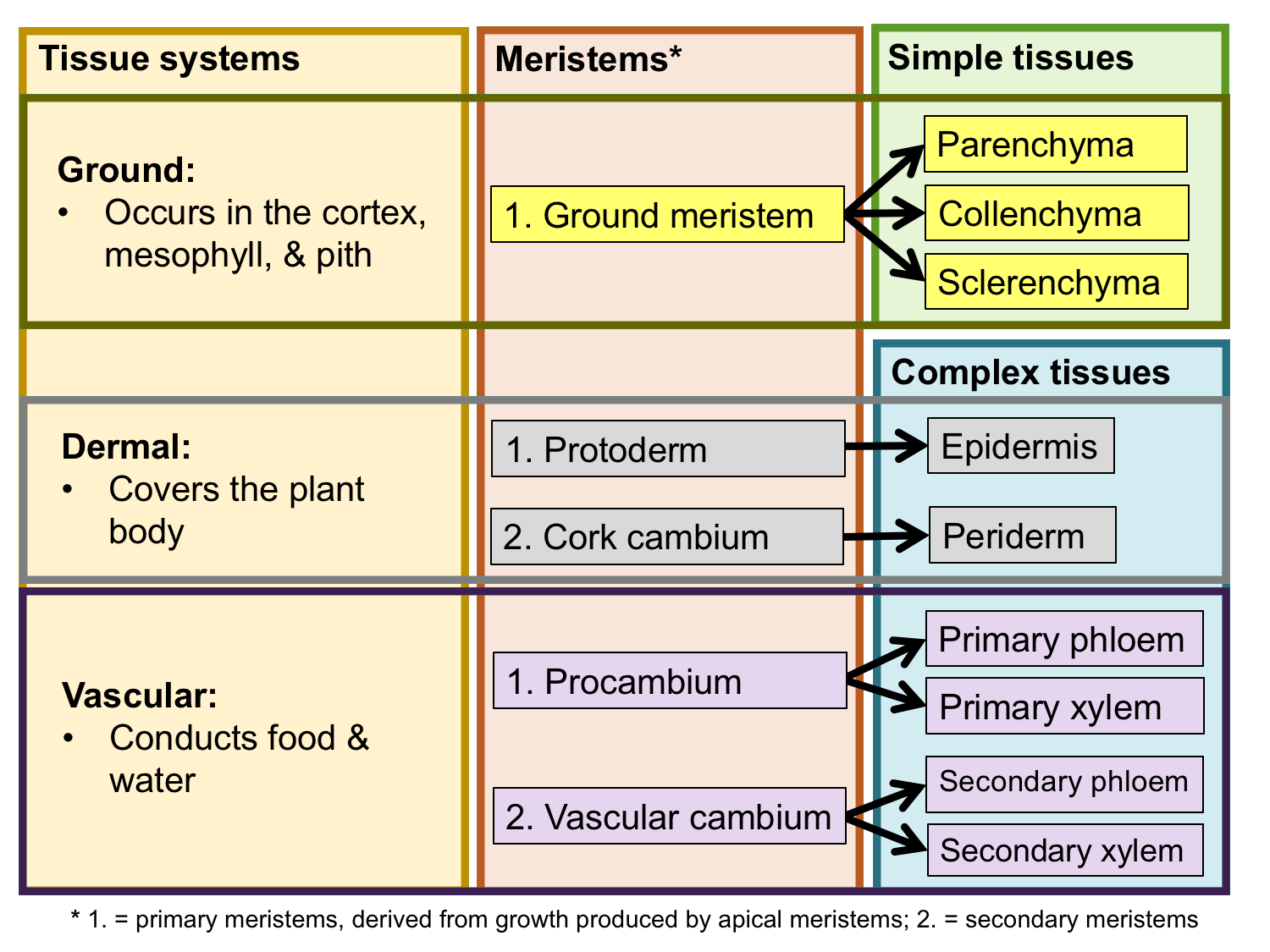
Tissue systems, meristems, and tissues. The chart above summarizes the relationship between the three tissue systems, the meristems that produce them, and the simple and complex tissues that comprise them. Credit: E.J. Hermsen (DEAL).
Selected references & further reading
Note: Free access is provided by the publisher for items marked with a green asterisk. Note that restrictions may apply for reuse.
Academic articles & book chapters
* Angiosperm Phylogeny Group, M.W. Chase, M.J.M. Christenhusz, M.F. Fay, J.W. Byng, W.S. Judd, D.E. Soltis, D.J. Mabberley, A.N. Sennikov, P.S. Soltis, and P.F. Stevens. 2016. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society 181: 1–20. https://doi.org/10.1111/boj.12385
* Chaw, S.-M., C.L. Parkinson, Y. Cheng, T.M. Vincent, and J.D. Palmer. 2000. Seed plant phylogeny inferred from all three plant genomes: Monophyly of extant gymnosperms and origin of Gnetales from conifers. PNAS 97: 4086–4091. https://doi.org/10.1073/pnas.97.8.4086
* Davis, C.C., Z. Xi, and S. Mathews. 2014. Plastid phylogenomics and green plant phylogeny: almost full circle but not quite there. BMC Biology 12, article number 11. https://doi.org/10.1186/1741-7007-12-11
* Doyle, J.A. 2013 [republished online 2018]. Phylogenetic analyses and morphological innovations in land plants. In: B.A. Ambrose and M. Purugganan, eds. The Evolution of Plant Form. Annual Plant Reviews 45. https://doi.org/10.1002/9781119312994.apr0486
* Pteridophyte Phylogeny Group [PPG I]. 2016. A community-derived classification for extant lycophytes and ferns. Journal of Systematics and Evolution 54: 563–603. https://doi.org/10.1111/jse.12229
* Wheeler, E.A. 2011. InsideWood - a web resource for hardwood anatomy. IAWA Journal 32: 199–211. https://doi.org/10.1163/22941932-90000051
* Xi, Z., J.S. Rest, and C.C. Davis. 2013. Phylogenomics and coalescent analyses resolve extant seed plant relationships. PLoS ONE 8(11): e80870. https://doi.org/10.1371/journal.pone.0080870
Books & textbooks
Esau, K. 1953. Plant Anatomy. John Wiley & Sons, New York.
Esau, K. 1977. Anatomy of Seed Plants, 2nd. ed. John Wiley & Sons, New York.
Evert, R.F., and S.E. Eichhorn. 2013. Raven Biology of Plants, 8th ed. W.H. Freeman and Co., New York, New York.
Peterson, R.L., C.A. Peterson, and L.H. Melville. 2008. Teaching Plant Anatomy through Creative Laboratory Exercises. NRC Research Press, Ottawa, Ontario, Canada.
Web resources: plant anatomy
Note: If a link does not work, try copying the URL and pasting it into your browser.
* Berkshire Community College Bioscience Image Library: Plant tissues and organs: http://blogs.berkshirecc.edu/bccoer/plants/
* Botanical Society of America Plant Image Collection: Plant anatomy: https://cms.botany.org/media/collection/id.24.html
* Cornell University Plant Anatomy Collection (CU-PAC): http://cupac.bh.cornell.edu/
* CU-PAC Plant Anatomy Online Lab Manual (M.A. Gandolfo, D.J. Paolillo, and J.L. Svitko): http://cupac.bh.cornell.edu/anatomy_manual/PAOnline.html
* InsideWood (2004 onwards, InsideWood Working Group and North Caroline State University): https://insidewood.lib.ncsu.edu/search
* Photographic Atlas of Plant Anatomy (J.D. Curtis, N.R. Lersten, and M.D. Nowak, revised by D. Hillier and J. Hardy): https://botweb.uwsp.edu/Anatomy/
* PhytoImages (contributed images of plant morphology and anatomy): http://www.phytoimages.siu.edu/
* Plant Anatomy Database (Department of Plant Biology, University of California, Davis; created by S. Detrick): https://anatomy.plb.ucdavis.edu/
* Plant Anatomy Laboratory: Micrographs of plant cells and tissues, with explanatory text (J.D. Mauseth): http://www.sbs.utexas.edu/mauseth/weblab/
* Plant Anatomy: Online Microscopy (Olivier Leroux): https://onlinemicroscopy.ugent.be/introENG.htm
* Science and Plants for Schools (SAPS) Image Collection: https://www.saps.org.uk/library/plant-science-image-library
* The Swingle Plant Anatomy Reference Collection (University of Miami): http://swingle.miami.edu/plantReference.html
* University of California Santa Barbara: Plant Anatomy (images by Vernon I. Cheadle and Katherine Esau): https://www.ccber.ucsb.edu/collections/botanical/plant_anatomy
* University of Wisconsin-Madison Botany Department Teaching Collection: https://uwdc.library.wisc.edu/collections/genbot/
Content usage
Usage of text and images created for DEAL: Text on this page was written by Elizabeth J. Hermsen. Original written content created by E.J. Hermsen for the Digital Encyclopedia of Ancient Life that appears on this page is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Original images and diagrams created by E.J. Hermsen are also licensed under Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
Content sourced from other websites: Attribution, source webpage, and licensing information or terms of use are indicated for images sourced from other websites in the figure caption below the relevant image. See original sources for further details. Attribution and source webpage are indicated for embedded videos. See original sources for terms of use. Reproduction of an image or video on this page does not imply endorsement by the author, creator, source website, publisher, and/or copyright holder.
Adapted images. Images that have been adapted or remixed for DEAL (e.g., labelled images, multipanel figures) are governed by the terms of the original image license(s) covering attribution, general reuse, and commercial reuse. DEAL places no further restrictions above or beyond those of the original creator(s) and/or copyright holder(s) on adapted images, although we ask that you credit DEAL if reusing an adapted image from the DEAL website. Please note that some DEAL figures may only be reused with permission of the creator(s) or copyright holder(s) of the original images. Consult the individual image credits for further details.
First released 30 March 2020; last updated 26 January 2021.