Chapter contents:
Class Bivalvia: Introduction and Morphology
– 1. Bivalve Phylogeny and Classification
– 2. Bivalve Ecology and Paleoecology
– 3. Evolutionary History of Bivalves ←
– 4. References and Further Reading
Image above: A fossil scallop (Pectinidae) from the Eocene Ocala Limestone of Florida. In this formation, mollusks with aragonitic shells are preserved only as casts and molds, while those with calcitic shells—such as this scallop—are unaltered.
Paleozoic Bivalves
The oldest known bivalves were tiny forms in the Early Cambrian. They are currently placed in their own order (Fordillida) without modern representatives or close relatives. Fordilla was one of these earliest bivalves, characterized by its small size, anterior adductor muscle larger than the posterior, and a single hinge tooth in each valve.

The bivalve Fordilla troyensis from the Early Cambrian of Rensselaer Co., NY (PRI 50316). Photo by Vicky Wang.
There are no known bivalve fossils between the Middle Cambrian and the Early Ordovician, when there was a major increase in body size and an explosion of forms into a variety of habits, including epifaunal and shallow infaunal species.
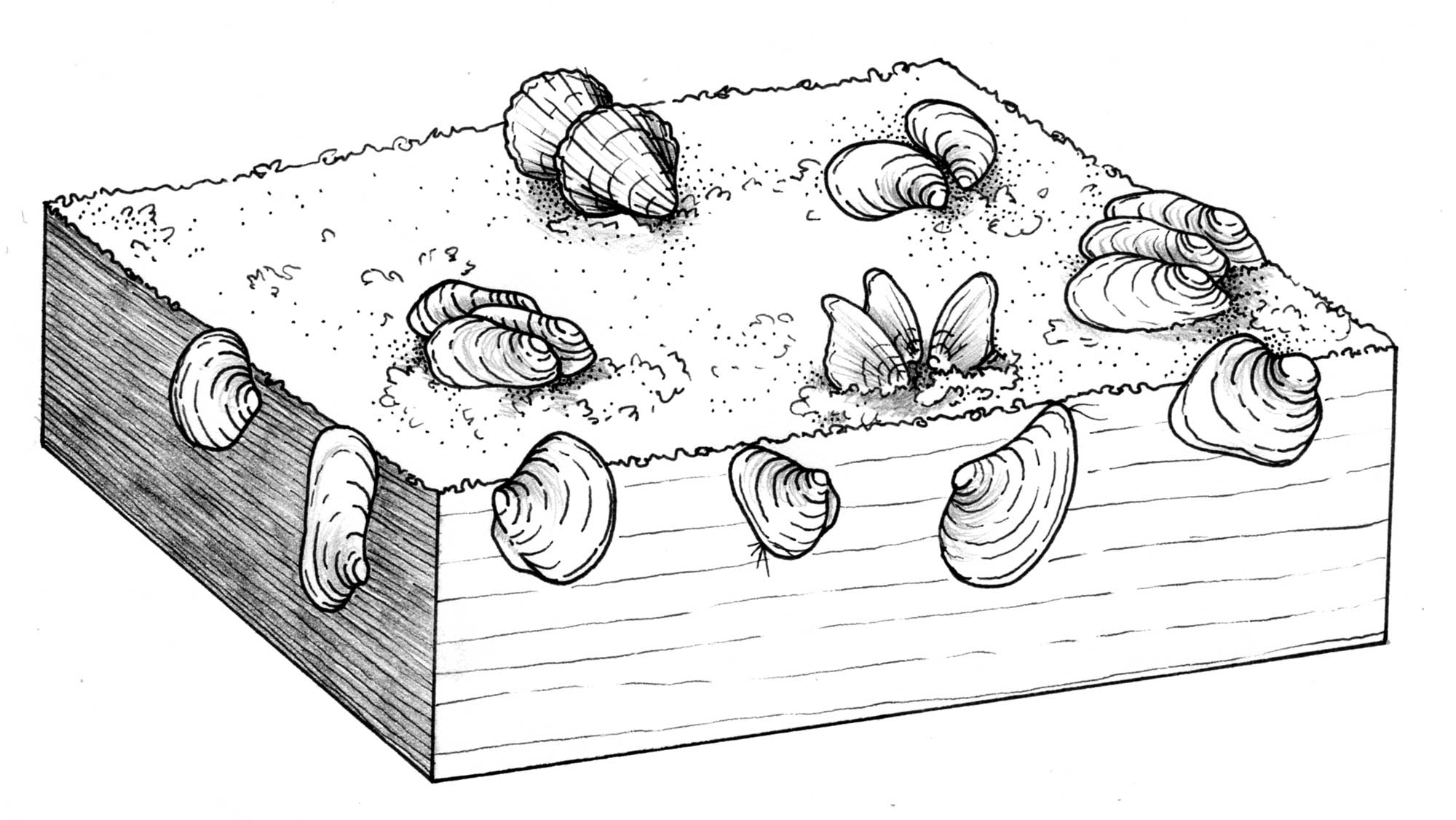
Reconstruction of the life habits of Ordovician bivalves. Drawing by Christi Sobel, based on Pojeta (1987).
Specimens of the Ordovician bivalve Ambonychia ulrichi from the Fairview Formation of Warren County, Ohio (PRI 49768). Specimen is on display at the Museum of the Earth, Ithaca, New York. Longest dimension of rock is approximately 13 cm (Sketchfab; Creative Commons 0 license).
Specimen of the Ordovician bivalve Vlasta americana (PRI 49892). Specimen is on display at the Museum of the Earth, Ithaca, New York. Longest dimension of shell is approximately 10 cm (Sketchfab; Creative Commons 0 license).
Byssal attachment first evolved in the Early Ordovician, and this led to changes in body symmetry and the anisomyarian and monomyarian conditions. This evolutionary diversification also apparently corresponded with the origin of the autobranch (W-shaped) gill. Bivalves maintained approximately this array of ecologies for the next 200 million years, during which time they were usually less diverse and abundant than brachiopods in the world’s oceans.
The oldest known freshwater bivalves are from the Devonian, but the major non-marine radiation occurred in the Triassic with the appearance of the Unionoida, which today includes more than 960 species, and is one of the most endangered animal groups on Earth due to habitat loss and specialized life cycle.

The Devonian freshwater bivalve Archanodon catskillensis. Left: Specimen from Lycoming County, PA (PRI 84851). Right: Specimen from Greene County, NY, in the collection of the Dept. of Geology, Franklin & Marshall College.

.jpg){kind=link}
Mesozoic Bivalves
The mass extinction at the end of the Permian dramatically changed the structure of life in the ocean. Brachiopods suffered much more severe extinction than did bivalves, which soon emerged in the Early Triassic as the dominant group of bottom-living marine invertebrates (learn more here).

Triassic marine bivalves from Blacktail Creek, Montana. Scalebars are 1cm unless otherwise denoted. a. Promyalina cf. spathi. b-c. Obnixia sp. d. Sp. A (infaunal bivalve species). e-f. Lingularia sp. g. Unionites canalensis. h. Unionites sp. i. Claraia extrema. j. Permophorus sp. k. Eumorphotis cf. multiformis. Courtesy of Elizabeth Petsios.
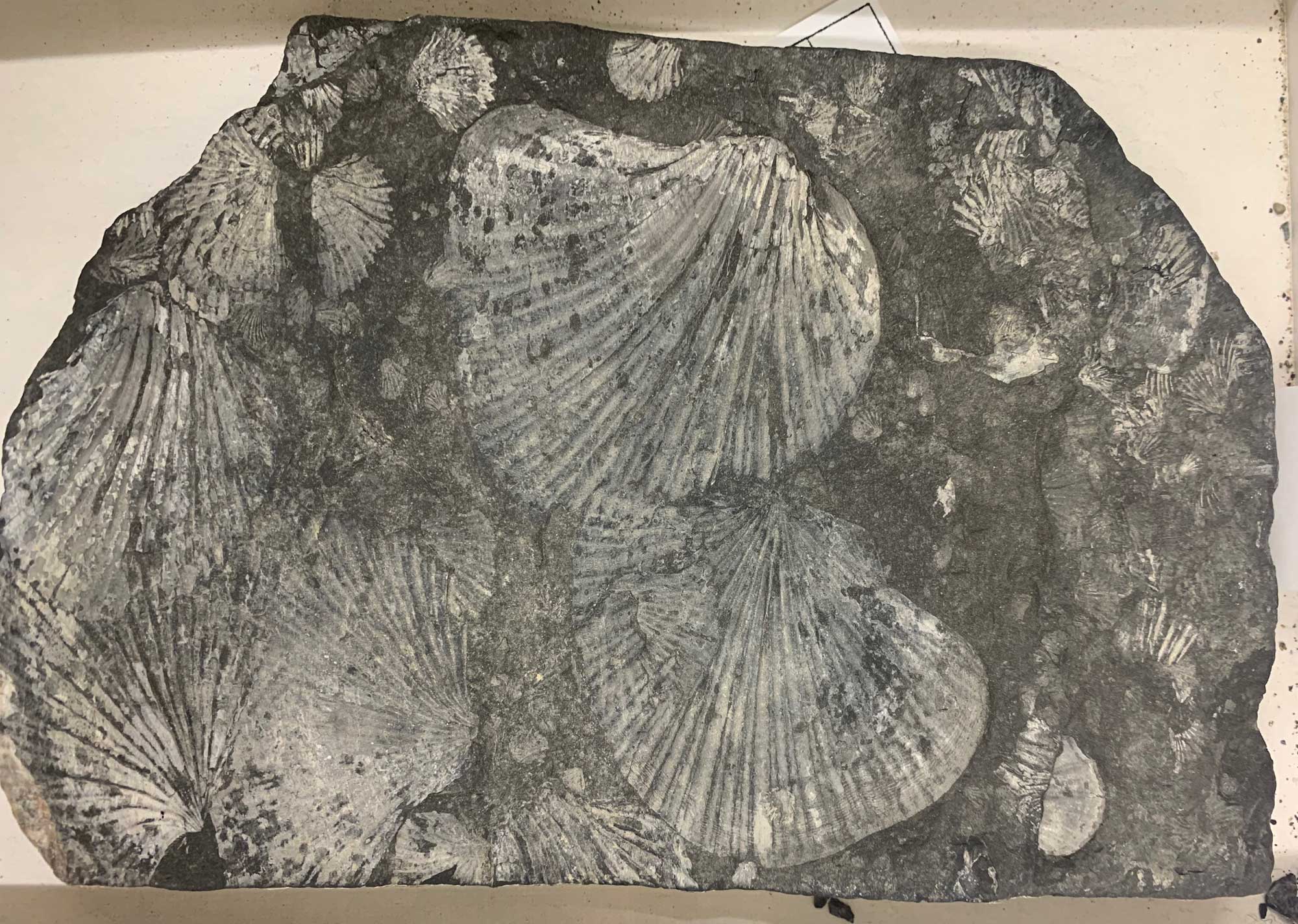
Specimens of the Triassic marine bivalve Monotis subcircularis from the Upper Triassic of British Columbia. Slab size is 15 x 20 cm (PRI Acc. 1307c).
During the Mesozoic, bivalves diversified dramatically in many marine habitats, from shallow epicontinental seas to deep-sea seeps. Cementation of one valve to the substrate evolved several times independently in the late Paleozoic and thereafter, including in the “true oysters” (Ostreidae) in the middle Triassic. One of the most important evolutionary innovations at this time was the fusion of the mantle edges to form siphons long enough to allow for deep burrowing in soft and hard substrates. This allowed bivalves to invade ecological niches that had never been available to brachiopods or most other phyla.
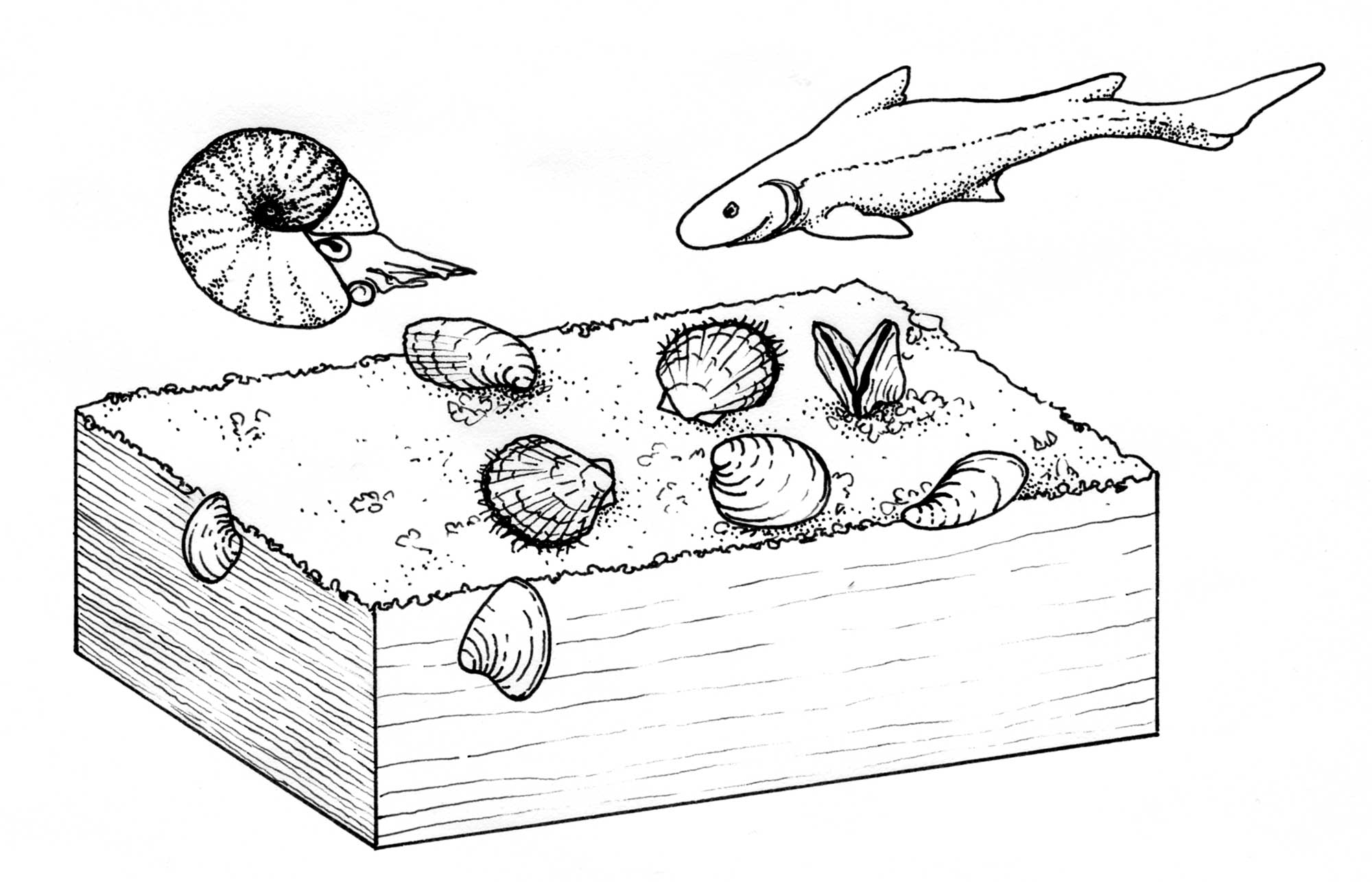
Reconstruction of marine bivalves and their habitat in the Early Triassic. Drawing by Christi Sobel, based on Chen and Benton (2012).
Mesozoic bivalves included some of the most unusual and distinctive fossil invertebrates, including inoceramids, rudists, and coiled oysters.
Inoceramids
Inoceramids were a group of pteriomorph bivalves that originated in the Permian but became extremely widespread, abundant, and diverse in the mid-late Cretaceous, when they are biostratigraphically very important. They were distinctive in frequently being the dominant epifauna (living on the surface of the sea floor) across wide areas of find-grained ocean bottoms, especially in organic-rich, low-oxygen environments. Inoceramids were frequently flat, and include some of the largest known bivalves, occasionally reaching 2-3 m in length.

Giant inoceramid bivalves. Left: Magadiceramus (?) rangatira rangatira from the Late Cretaceous (latest Cenomanian) of Rout Point, Kaipara Harbour, Auckland, New Zealand (Fossil Record Number Q08/f328), Institute of Geological and Nuclear Sciences (GNS Science), Wellington, New Zealand; photograph by Warren D. Allmon. Middle: Sphenoceramus/Inoceramus steenstrupi from Greenland on exhibit in the Zoological Museum of Copenhagen, maximum length 1.87 m; image by Mike Beauregard (Wikimedia Commons; Creative Commons Attribution 2.0 Generic license; image resized and incorporated into plate shown above). Right: A large specimen of the inoceramid Platyceramus platinus from the Late Cretaceous Niobrara Chalk of Kansas on display at the Denver Museum of Science and Nature (longest dimension is approximately 1.2 m in length); photograph by Jonathan R. Hendricks.
Their shells were distinctive for having a very prominent prismatic layer (which frequently falls apart into separate prisms of calcite after death), a single adductor muscle (monomyarian), prominent growth lines that appear as raised concentric semicircles, and weak or no hinge teeth. The presence of chemosymbiotic bacteria in their tissues may have allowed them to colonize such inhospitable habitats.
Rudists
Rudists were a group of highly derived heterodont bivalves with one or both valves taking on an elongated cylindrical form (in addition to the 3D model below, see photos on previous pages in this chapter).
The rudist bivalve Sellaea sp. from the Cretaceous Edwards Limestone of Hill County, Texas (PRI 42055). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Length of specimen is approximately 9 cm.
They were diverse and abundant from the Late Jurassic to the end of the Cretaceous, and became the most important reef-formers of this time. Rudists varied from uncemented, doubly-coiled species that “reclined” on the sea floor, to upright and cemented forms with one enormously elongated valve and another cap-shaped valve.

Reconstruction of a group of coiled Cretaceous rudist bivalves. Image copyrighted by the Smithsonian Institution (SI) and shared here in accordance with the SI terms of use for educational and noncommercial purposes.
Coiled Oysters
Coiled oysters, such as Exogyra and Gryphaea, were diverse and abundant during the Jurassic and Cretaceous.
Their unusual morphology apparently suited them to live epifaunally on both soft and firm substrates. They may have declined in diversity after the Mesozoic because of increased predation and bioturbation associated with the Marine Mesozoic Revolution.
Cenozoic Bivalves
The mass extinction at the end of the Cretaceous included the loss of approximately 65% of bivalve genera and possibly up to 80% of species. Epifaunal species diversity declined relative to infaunal species. From the survivors, diversity quickly increased in the Paleogene and continued to expand through the rest of the Cenozoic.

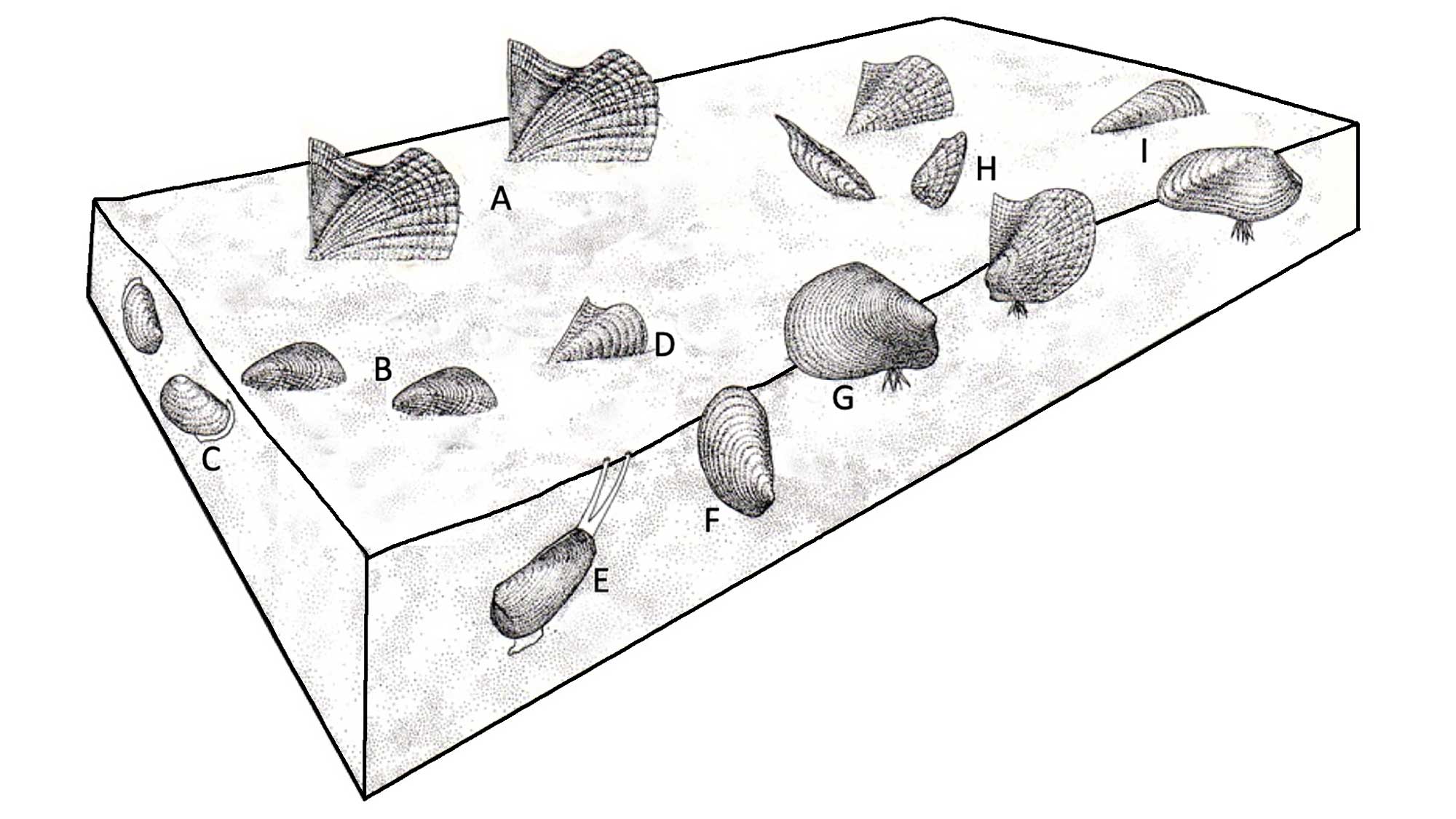
Evolutionary radiation of infaunal bivalves following evolution of mantle fusion in the late Paleozoic/early Mesozoic; modified from Stanley (1968).
Today both infaunal and epifaunal marine bivalves show a strong latitudinal diversity gradient, with the only exception to this being the deposit-feeding protobranchs. Although a latitudinal diversity gradient probably existed as long ago as the late Paleozoic, it steepened significantly since the mid-Mesozoic, mainly because of major diversification of infaunal heterodonts (Imparidentia).
Shells of the bivalve Eurhomalea sp. (Imparidentia) from the Eocene La Meseta Formation of Antarctica (PRI 65454). Specimen is from the research collections of the Paleontological Research Institution, Ithaca, New York. Longest dimension of specimen is approximately 13.5 cm (Sketchfab; Creative Commons 0 license).
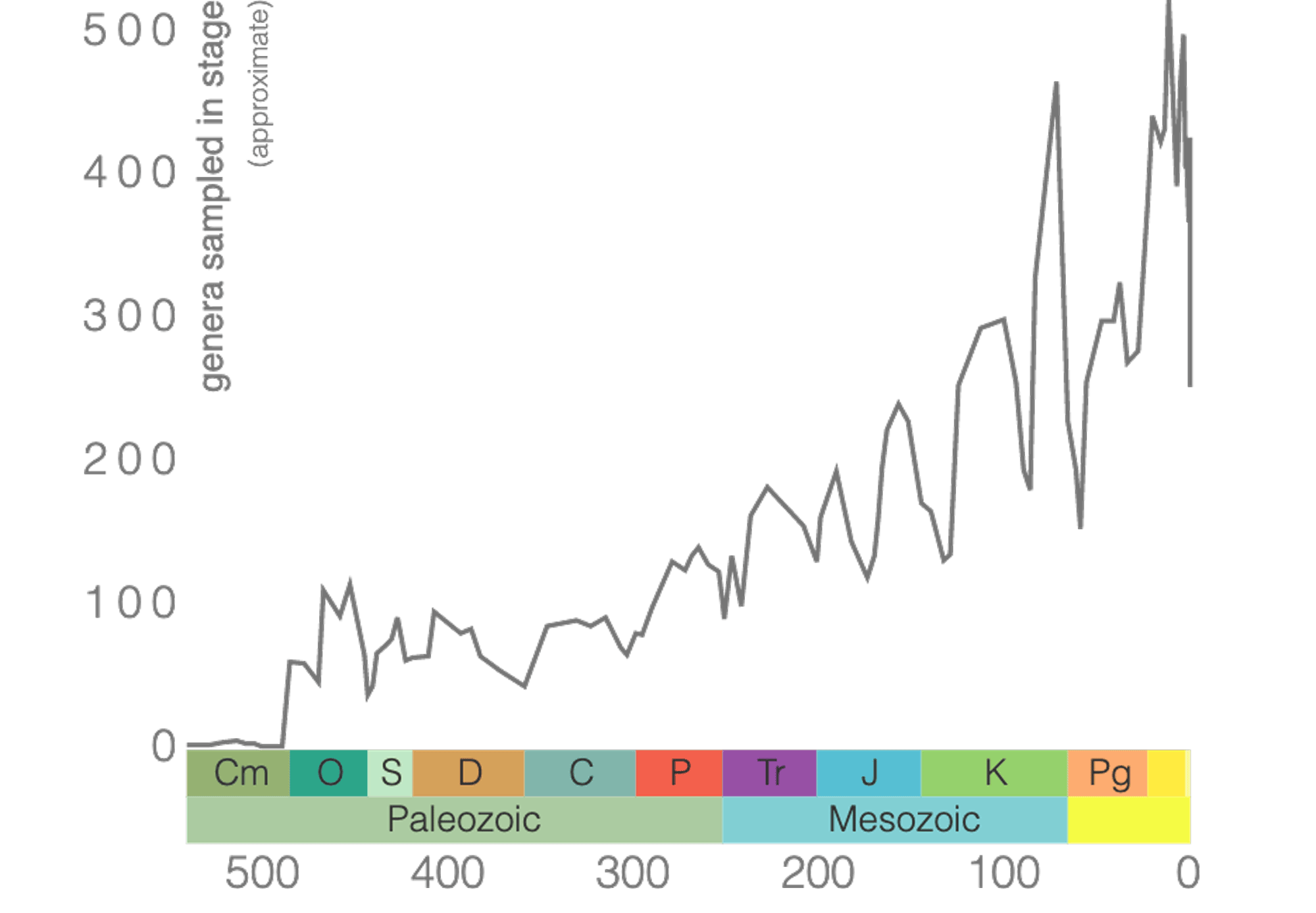
Phanerozoic Diversity of Bivalves
Bivalve diversity increased quickly in the Ordovician but then plateaued for most of the Paleozoic Era. Bivalves were only slightly affected by the end-Permian mass extinction, and soon began an irregular increase in diversity through the Mesozoic. They suffered significant diversity decline at the end-Cretaceous mass extinction, but have increased in diversity almost continuously throughout the Cenozoic.