


Chapter contents:
Class Gastropoda: Introduction, Morphology, Anatomy, and Life History
– 1. Phylogeny and Classification of Extant Gastropoda ←
– 2. Fossil Record of Gastropoda
– 3. References and Further Reading
– 4. Silly Snail Stuff
Image above: Modern strombid conch shells from Sanibel Island, Florida; collections of the Paleontological Research Institution.
Overview
Scientific understanding of the evolutionary relationships among gastropods has changed dramatically over the past several decades. Gastropod classification has, therefore, been in a long period of instability. In the last few years, however, something of a consensus has emerged about the major features of gastropod phylogeny, and a stable classification is now developing.
For most of the twentieth century, gastropods were divided into three major groups (frequently referred to as “subclasses”): Prosobranchia, Opisthobranchia, and Pulmonata. Opisthobranchs and pulmonates were frequently grouped together as Euthyneura. Prosobranchia was also traditionally divided into three groups: Archaeogastropoda, Mesogastropoda, and Neogastropoda. Starting in the 1980s, however, studies of microanatomy and genetics indicated that most of these were not monophyletic groups, and that new hypotheses and arrangements were needed.
Gastropod evolutionary relationships are difficult to unravel for several reasons. First, in studies of living species, molecular, anatomical, and shell form frequently support conflicting phylogenetic hypotheses. Second, gastropod shells—which have historically been the most-studied aspects of these animals—frequently have relatively few discrete characters that are amenable to phylogenetic analysis. This means that many fossil gastropods, especially from the Paleozoic (see Fossil Record of Gastropoda page), are very difficult to place into living groups, and it is frequently challenging to do phylogenetic analyses with fossil snail shells. Complicating matters further, the shells of gastropods are often shaped by adaptation, for example, to provide protection from predators, and may therefore be subject to significant evolutionary convergence, resulting in homoplasy in phylogenetic reconstructions.
It is now clear that similar shell shapes have evolved in gastropods multiple times independently. For example, cap-like shells have evolved multiple times in Gastropoda that are commonly known as "limpets." Thus, “limpets” do not constitute a single monophyletic group, but rather are polyphyletic in origin. Many other shell forms have similarly evolved independently multiple times, for example shells with very high spires, shoulders, spines, or overall shapes.

Convergent evolution (homoplasy) in gastropod shells; note that shells are not shown at the same scale. Figs. 1-3. The shells in figs. 1-3 share an elongate, high-spired shape, but fig. 1 (Terebra aclinica, Pliocene of Florida, PRI 41846) is a neogastropod in the family Terebridae, whereas the shells in fig. 2 (Turritella praecincta, Paleocene of Alabama, Museum of Comparative Zoology, Harvard University MCZIP 29349) and fig. 3 (Palmerella mortoni postmortoni, Paleocene of Alabama, PRI collection) are cerithioids in the family Turritellidae. The two turritellids (figs. 2, 3) also are convergent in having sharp spiral ridges, or carinae, but in fig. 2 the carina is at the top of each whorl (just below the suture) while in fig. 3 it is at the base (far below the suture). Figs. 4, 5. The shells in figs. 4 and 5 both show prominent shoulders on the body and spire whorls, but the specimen in fig. 4 (Bullia (Bulliopsis) quadrata, Miocene of Maryland, PRI 104728) is a neogastropod in the family Nassariidae, whereas fig. 5 is a cerithioid (Melanopsis martini, Pliocene of Europe, PRI 42207) in the family Melanopsidae. Very high-spired shells may function to allow the animal to pull its body far inside to avoid predation, but the function (if any) of carinae and shoulders is unknown. Figs. 6, 7. The specimens in figs. 6 and 7 have very similar overall shapes, but fig. 6 is a cone snail (Conus anabathrum, Plio-Pleistocene of Florida; PRI 54676) from the family Conidae, whereas the shell in fig. 7 is a dove snail (Parametaria islahispaniolae, Miocene of the Dominican Republic) from the family Columbellidae. Note that the dove snail has denticles (tooth-like bumps) on the inside of the lip, a feature that is never present in cone snails.
Similarly, "land snails" independently invaded the land from the sea at least 12 times over their evolutionary history (Ponder and Lindberg, 2020), so also represent a polyphyletic grouping. Of the many independent gastropod groups that have transitioned to land, those in the Stylommatophora (Heterobranchia) and Caenogastropoda clades are the most diverse today, with 20,000 and 4,200 species respectively (Ponder and Lindberg, 2020). Although land snails can be locally abundant as fossils, many extant land snail groups have a poor or non-existent fossil record.

Examples of modern land snail shells from the collections of the Paleontological Research Institution. Photograph by Jonathan R. Hendricks
The history of gastropod classification is filled with many terms referring to different taxonomic groups, which can be very confusing. These terms have not always been used consistently, and many older and obsolete terms are still used by biologists and paleontologists. The following six major clades of living gastropods are now generally accepted and each is elaborated upon below:
A recent phylogenetic hypothesis for these major clades, developed by Uribe et al. (2019), is shown below. Note that Caenogastropoda and Heterobranchia form a clade that is known as Apogastropoda. Collectively, Apogastropoda + Neritimorpha + Vetigastropoda + Neomphalina (i.e., all extant gastropoda except Patellogastropoda) form a clade called Orthogastropoda. Patellogastropoda, the true limpets, is the sister group of Orthogastropoda.
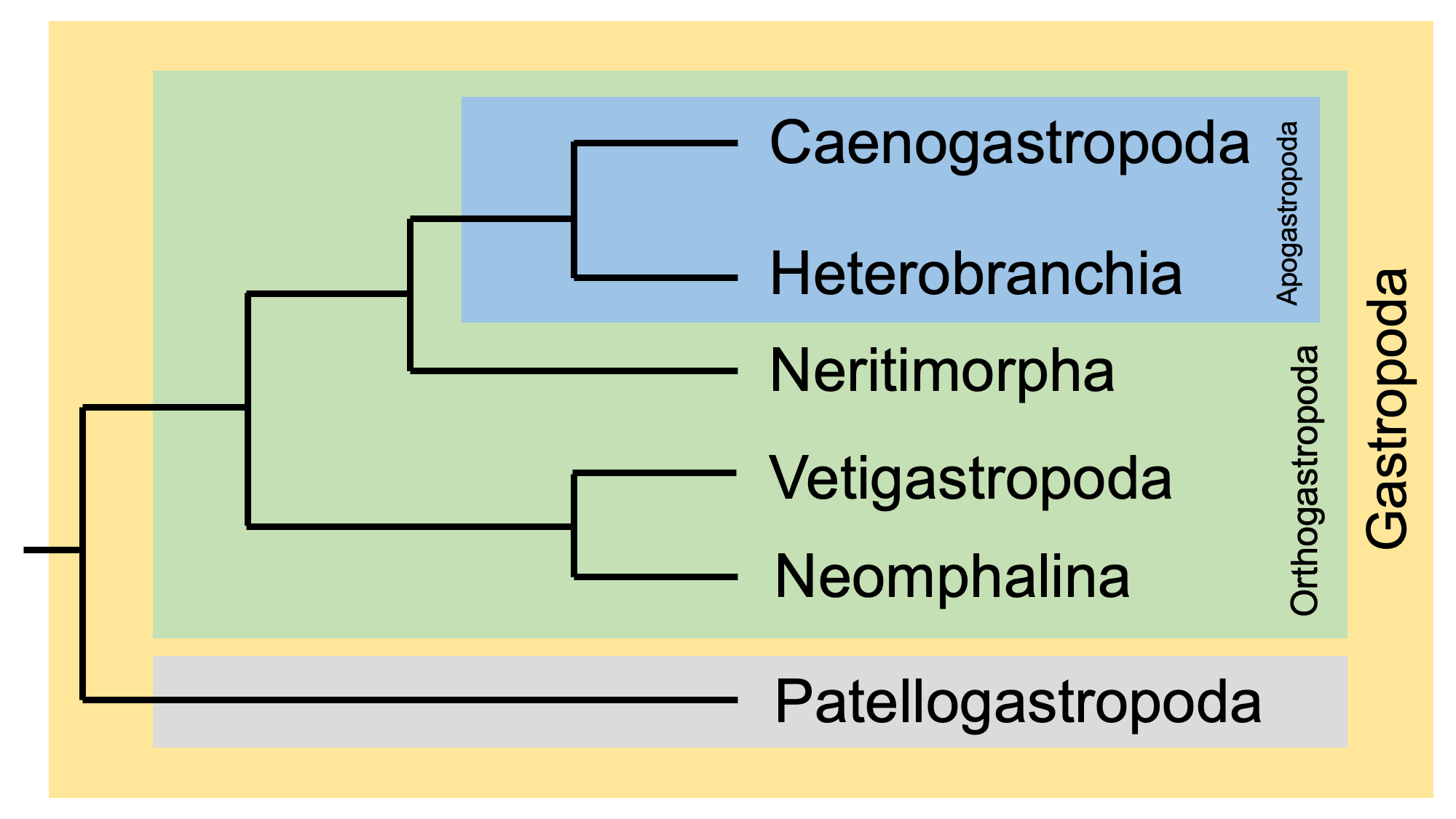
Phylogenetic hypothesis for the six major clades of extant gastropods. Redrawn and simplified from Uribe et al. (2019, fig. 5). Image by Jonathan R. Hendricks for the Digital Atlas of Ancient Life Project.
Alternatively, a different recent study by Cunha and Giribet (2019) found Vetigastropoda and Patellogastropoda to be sister taxa with one another, and in turn sister taxa to the other three major gastropod clades (these authors did not include Neomphalina in their analysis). The relationships between major gastropod clades thus remain a matter of debate.
Extant Gastropod Clades
Quick links to clades:
Subclass Caenogastropoda
Overview
Caenogastropoda is the most diverse clade of gastropods, including about 157 extant families, 190 extinct families (Bouchet et al., 2017), and 60% of living snail species (>23,000; WoRMS). They are incredibly varied in morphology, ecology, and behavior. Most are marine, but others live in freshwater and on land. Some eat algae, while others use venom to hunt prey, including fish. Most caenogastropods have a shell, but it never has a nacreous layer. There is a single gill (ctenidium). Living species have either planktotrophic or (more rarely) non-planktotrophic larvae, with orthostrophic protoconchs.
Name origins: “Caeno-” is a variant of “Ceno-,” which is Greek for new. Thus, caenogastropods are “new” or more recent gastropods.
Until recently, caenogastropods were considered to be within “prosobranchs,” including most of the “mesogastropods” and all of the “stenoglossans” or neogastopods. Phylogenetic relationships within caenogastropods are still topics of debate, with internal relationships changing frequently. The image below shows a recent phylogenetic hypothesis for Caenogastropoda simplified from the molecular phylogeny of Irwin et al. (2021).
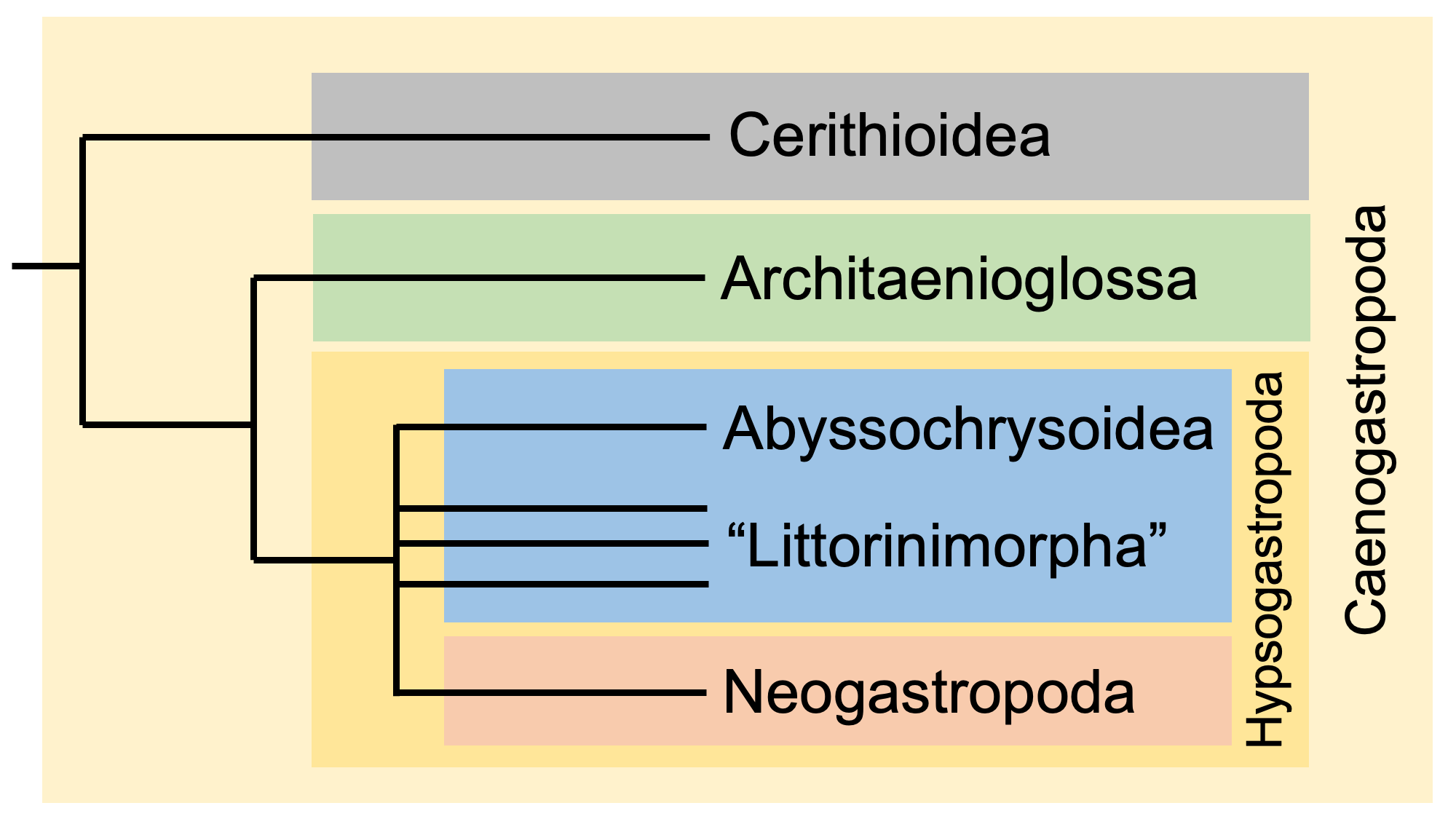
In this hypothesis, Cerithioidea is the basal-most clade. Architaenioglossa, which includes a number of freshwater and terrestrial taxa, forms the next branch up from the base. Hypsogastropoda is the largest clade of Caenogastropoda and includes several groups. "Littorinimorpha" is a paraphyletic group comprised of hypsogastropods that lack a siphonal canal; our understanding of the relationships among littorinimorphs is in a nearly constant state of flux. Abyssochrysoidea is a small clade composed of deep water species that inhabit hydrothermal vent systems. The largest clade of hypsogastropods—and the largest clade of gastropods—is Neogastropoda, a group that is nearly always recovered as monophyletic in analyses and is comprised of carnivorous species. Neogastropods all have shells with a siphonal canal, which holds an inhalant siphon through which water enters the mantle cavity, and an elongate proboscis, which is often used to seize prey.

The siphon of a Venus comb murex (Murex pecten) extending from the shell's siphonal canal. Photograph by Samuel Chow (Flickr; Creative Commons Attribution 2.0 Generic license; image cropped and labels added).
Neogastropods include some of the most derived gastropods, such as the cone snails (see below), which use powerful neurotoxins in conjunction with harpoon-like delivery systems to immobilize and capture their prey. Phylogenetic relationships within Neogastropoda are still a topic of disagreement among researchers.
While numerous anatomical and molecular characters unite living caenogastropods, unequivocal identification of caenogastropods by shell characters alone is much more difficult. All have an orthostrophic protoconch, but this delicate feature is often not preserved in fossils. Orthostrophic protoconchs are also present in Neritimorpha, but they otherwise have very different teleoconchs. The oldest undoubted caenogastropods are from the Devonian. The group, especially the basal Cerithiimorpha, began to diversity in the late Paleozoic, and this accelerated greatly in the Mesozoic.
Detailed taxonomic information about the subgroupings of extant Caenogastropoda may be found on MolluscaBase.
Examples of Caenogastropoda
Cigarette Cone: Conus Geographus

The Indo-Pacific cone snail Conus geographus is sometimes referred to as the “cigarette cone” for an ominous reason: if you are stung by this neogastropod, you may only live long enough to smoke a single cigarette before you die. Like all cone snails, Conus geographus is a venomous predator. Unlike most cone snails—a majority of which eat worms—the cigarette cone hunts small fish. Because its neurotoxins, termed “conotoxins,” target the vertebrate nervous system (many aspects of which humans and fish share in common), defensive stings delivered to unwary shell collectors can be fatal. University of Washington Professor Emeritus Alan Kohn has spent much of his career researching the natural history of cone snails and was the first to discover that this particular species eats fish. Dr. Kohn recently reviewed all medically documented cases of cone snail stings and found that Conus geographus has been responsible for at least 30 human deaths (Kohn, 2016), which is typically caused by muscle paralysis leading to respiratory failure. An envenomation event in Queensland, Australia in 2015–perhaps by Conus geographus–is recounted here. For more information about the “cigarette cone”–a type of neogastropod–watch the video below.
Queen Conch: Lobatus gigas

Specimens of the Queen Conch, Lobatus gigas in the collections of the Paleontological Research Institution. Photograph by Jonathan R. Hendricks for the Digital Atlas of Ancient Life project.
The large shells of the Queen Conch, Lobatus gigas, are perhaps the most iconic of all Caribbean seashells. The flesh of this herbivorous species is edible, but over harvesting has resulted in dwindling populations. The video below reviews conservation efforts associated with the Queen Conch, which belongs in the Stromboidea clade (traditionally classified as a group of Littorinimorpha).
Apple Snails: Pomacea

Modern Pomacea paludosa, from Lake Tsala Apopka, Florida (PRI 70096). The hole in the dorsum of the shell was likely made by the curved bill of a Snail Kite (Rostrhamus sociabilis), a bird of prey that feeds almost exclusively on this species of snail. Image by Jonathan R. Hendricks for the Digital Atlas of Ancient Life.
Pomacea is a genus of freshwater snails–commonly known as “apple snails” or “mystery snails”– that includes a species native to Florida. They belong to the Ampullaroidea clade and are traditionally classified as members of the Architaenioglossa. These snails are sometimes kept as pets and care tips are presented in the video below.
Angle-mouth Screw Shell: Turritella gonostoma

Specimens of Turritella gonostoma from San Carlos Bay, Guaymas, Mexico; collections of the Paleontological Research Institution. Photograph by Jonathan R. Hendricks for the Digital Atlas of Ancient Life project.

Left: A living individual of Turritella gonostoma, filtering food from the water. Right: Clusters of Turritella gonostoma eggs. Photographs by Warren D. Allmon.
Gallery of Caenogastropoda

Fossil Sinistrofulgur contrarium (Family Busyconidae) from the upper Pliocene Tamiami Formation (Pinecrest Beds) of Florida (PRI 70051).

Fossil Ficus sp. (Family Ficidae) from the upper Pliocene Tamiami Formation (Pinecrest Beds) of Florida (PRI 70047).

Fossil Strombus sp. (Family Strombidae) from the upper Pliocene Tamiami Formation (Pinecrest Beds) of Florida (PRI 70052).

Fossil Turritella magnasulcus (Family Turritellidae) from the upper Pliocene Tamiami Fm. (Pinecrest Beds) of Florida (PRI 70049).

Fossil Crucibulum auricula (Family Calyptraeidae) from the upper Pliocene Tamiami Fm. (Pinecrest Beds) of Florida (PRI 70055).

Fossil Calyptraea centralis (Family Calyptraeidae) from the upper Pliocene Tamiami Fm. (Pinecrest Beds) of Florida (PRI 70046).

Fossil Cymatosyrinx aclinica (Family Drillidae) from the upper Pliocene Tamiami Fm. (Pinecrest Beds) of Florida (PRI 70050).

Fossil Naticarius plicatella (Family Naticidae) from the upper Pliocene Tamiami Fm. (Pinecrest Beds) of Florida (PRI 70044).
Subclass Heterobranchia
Overview
Heterobranchia (also sometimes called Heterostropha) is a large and important group that includes two major subgroups: 1) the bubbleshells and shell-less nudibranchs (or, “sea slugs”) (previously known as “opisthobranchs”); and 2) most of the land snails and all terrestrial slugs (formerly known as “pulmonates”). Several smaller freshwater and marine groups are also included in Heterobranchia. The shells of heterobranchs always lack nacre, but are diverse in form. In some slugs, the shell is either absent or is greatly reduced in size and internalized. Some heterobranchs have limpet-shaped shells, a form that has evolved numerous times in gastropods. Most heterobranchs lack opercula. All types of larval developmental mode are represented among heterobranchs. Their protoconchs are typically sinistral, but adult shells exhibit heterostrophy. Heterobranchs do not have ctenidia, but one or more secondary gills are present and terrestrial species have vascularized mantle cavities that are modified to function as a lung.
Recent understanding of heterobranch relationships indicates the following major groups:
- “Lower Heterobranchia” (aka “Allogastropoda”): A paraphyletic assortment of predominantly marine gastropods that are phylogenetically basal. Example: the sundial shells, Architectonica.
- Euthyneura: A mostly monophyletic clade that includes three major subclades:
- Nudipleura: The marine nudibranchs, one group of sea slugs. Species often exhibit dazzling colors and fascinating defenses, including the ability to steal and co-opt the stinging cells (nematocysts) of some cnidarians. They lack shells as adults.
- Euopisthobranchia: Includes the opisthobranchs other than nudibranchs; marine sea slugs, many of which have shells. Some examples include sea hares, headshield slugs, bubble snails, and pteropods.
- Panpulmonata: Includes Sacoglossa, Pyramidellidae, and the traditional pulmonates; this clade contains mostly air-breathing, terrestrial gastropods, including common garden snails and slugs. Also includes two clades of air-breathing “false limpets,” Siphonarioidea and Trimusculoidea.
The oldest probable fossil heterobranchs (as indicated by a heterostrophic protoconch) occur in the Middle Devonian (Fryda et al, 2008: p. 257).
Name origins: “Hetero-,” different; “branchia,” gills. Thus, heterobranchs are “differently gilled” gastropods.
Examples of Heterobranchia
The Blue Dragon: Glaucus atlanticus
"The Real Life Sea Dragon" by ZoneA (YouTube).
The blue dragon, Glaucus atlanticus, is a planktonic, brightly-colored marine nudibranch (Nudipleura clade). It can weakly paddle, but cannot swim against a current. A 2010 viral video characterized this species as a “real life Pokemon.” This heterobranch protects itself by stealing and co-opting stinging cells from the Portuguese man-of-war; many nudibranchs similarly steal the weaponry of cnidarians for their own defense.
Miniature Melo Bubble Snail: Micromelo undatus
Bubble snails are euopisthobranchs. They crawl on the seafloor, on algae, or below the sand surface. Their bodies are usually larger than their shells, so some cannot retract completely into their shells. The video above shows a kind of Indonesian bubble snail called a Miniature Melo (Micromelo undatus).
California Aglaja Headshield Slug: Navanax inermis
Species belonging to the marine headshield sea slug genus Navanax are predatory members of the Euopisthobranchia clade. The short video above shows a Navanax inermis eating a a small sea hare (another kind of sea slug) by sucking it in whole.
Sea Butterflies and Sea Angels
Sea butterflies and sea angles are pteropods (“wing-foot;” the “p” is silent), both shown in the video above. These small euopisthobranchs spend their lives in the water column (i.e., they are pelagic). The fine calcium carbonate shells of sea butterflies—which easily dissolve in acidic water—have proven to be important indicators that the oceans are becoming more and more acidic due to the presence of higher amounts of dissolved carbon dioxide (see overviews by the Smithsonian and Los Angeles Times).
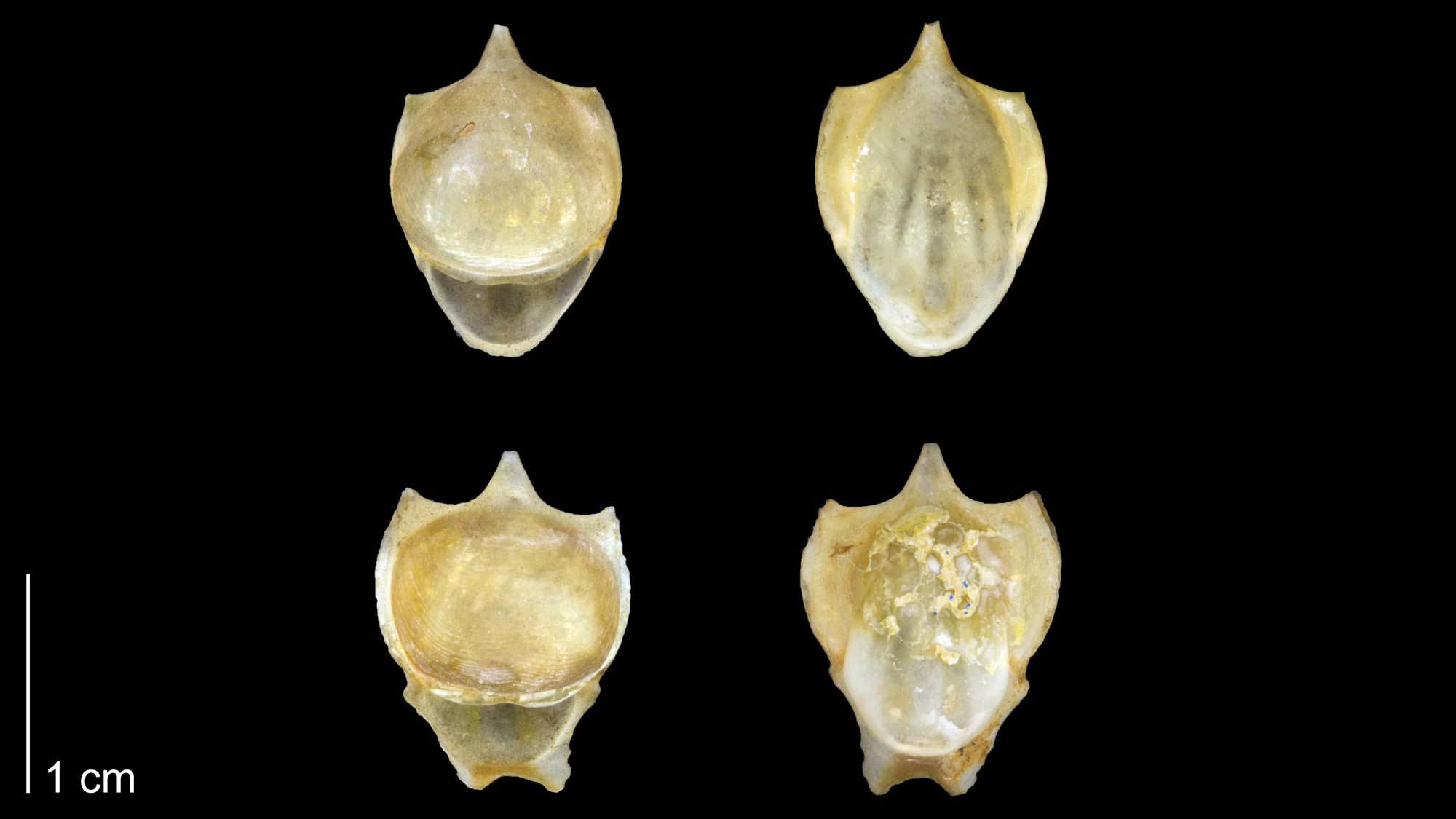
Shells of the extant pteropod gastropod Cavolinia tridentata , a type of sea butterfly (PRI 70106). Specimens are from Zanzibar and are held at the Paleontological Research Institution, Ithaca, NY. Image by Jonathan R. Hendricks for the Digital Atlas of Ancient Life project.
Brown Garden Snail: Cornu aspersum
Banana Slug: Ariolimax
"Banana Slugs: Secret of the Slime" by PBS Deep Look (YouTube).
Banana slugs (genus Ariolimax), along with other terrestrial slugs, are panpulmonates. These slimy, brightly colored slugs occupy the coastal forests of California, Oregon, and Washington.
Gallery of Heterobranchia

Modern specimen of the sundial snail Architectonica maxima ("Lower Heterobranchia") from Tosa, Japan (PRI 55220).

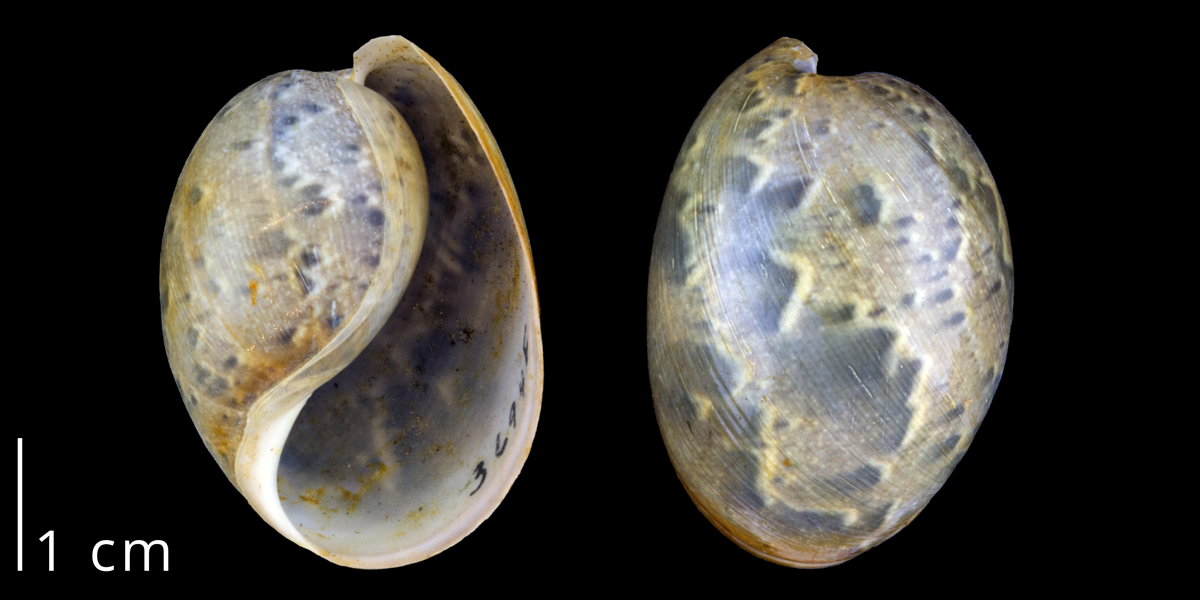
Shell of the modern bubble snail Bullaria gouldiana (Euopisthobranchia) from Orange County, California (PRI 70098).
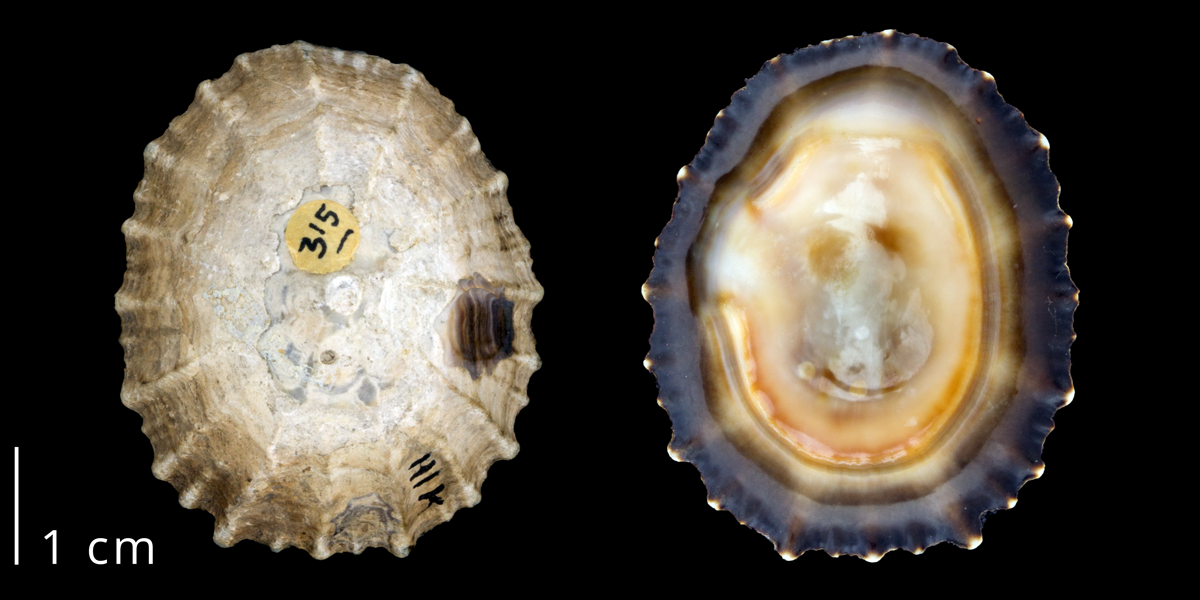
Modern shell of the false limpet Siphonaria gigas (Siphonariidae) from Panama (PRI 70100).
Subclass Neritimorpha
Overview
This group (also called Neritopsina) includes species previously included in the “Archaeogastropoda,” and generally shows a combination of primitive and derived features that has led to much uncertainty about their phylogenetic position. While less diverse than Caenogastropoda and Heterobranchia, the clade has ~2,000 extant species (more than 500 of which are in the terrestrial family Helicinidae) and is also ecologically varied, with members occupying marine, freshwater, and terrestrial habitats (Uribe et al., 2016). Similar to both of these larger clades, slug- and limpet-like forms have also independently evolved within the Neritimorpha (Uribe et al., 2016). Neritimorphs usually have small shells that are never nacreous, but vary in form from coiled to limpet-shaped. Many have shell apertures with folds or teeth on the inner lip. Most living species resorb the early whorls and columella as the animal grows. The shells of neritimorphs usually have an outer layer of homogeneous calcite and inner layers of cross-lamellar or prismatic aragonite. The veliger larvae are frequently planktotrophic with orthostrophic protoconchs. Neritimorphs have opercula and a single ctenidium.
Some marine genera of Neritimorpha are associated with particular environments; for example, Smaragdia occurs in association with seagrass, while Neritina lives in brackish water environments (Costa et al., 2001). Finding these same taxa in the fossil record therefore may be useful for interpreting past environments.
{kind=link}

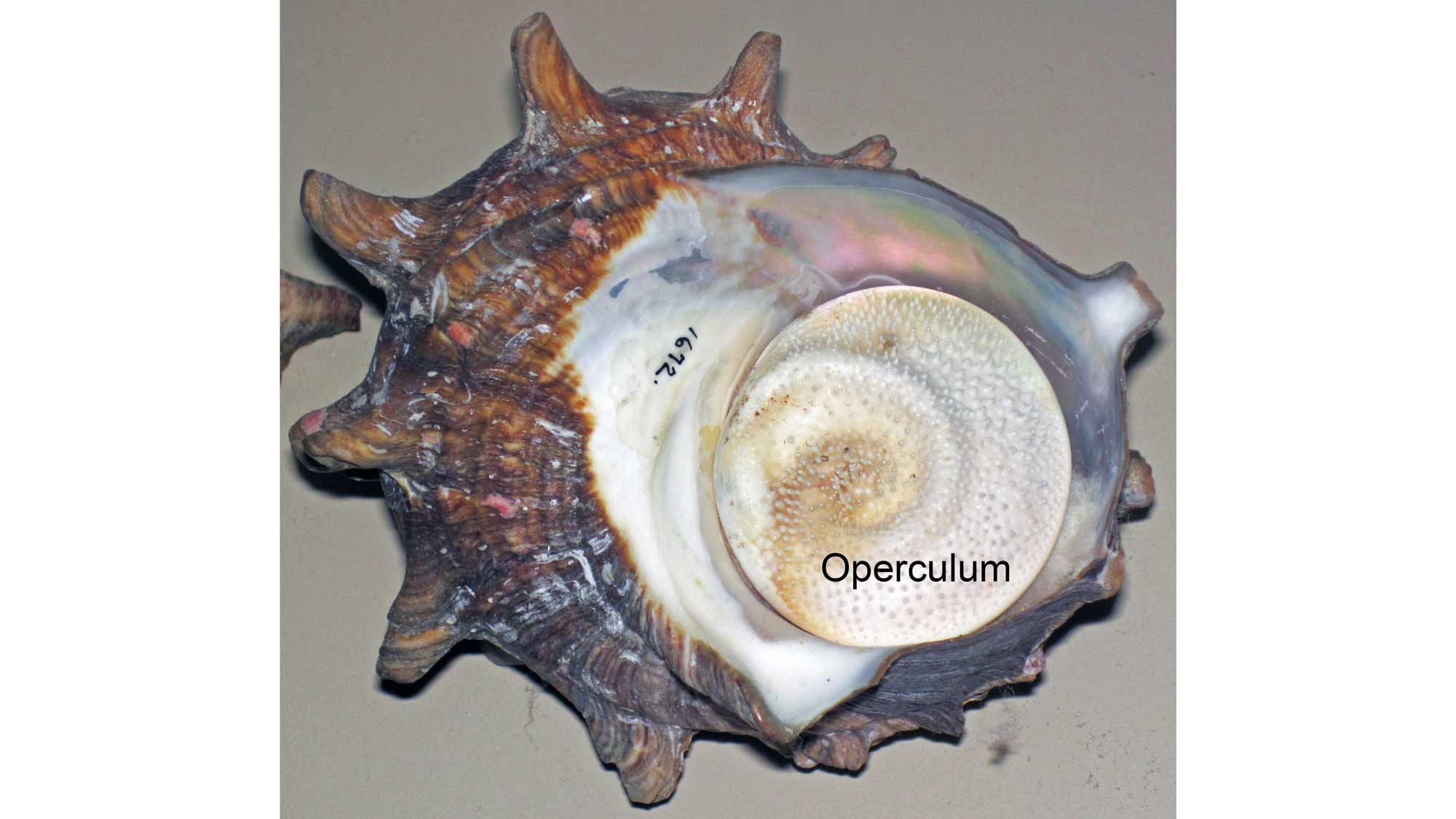
Modern bleeding tooth snail Nerita peloronta from Bermuda (PRI 70104). Note that the operculum of the shell is in situ (in place). Image by Jonathan R. Hendricks for the Digital Atlas of Ancient Life project.
Possible neritimorphs occur in the Ordovician although the oldest likely records (based on protoconch morphology) date to the Devonian or possibly Silurian. Neritimorphs may have evolved terrestrial forms at least three times during their history.
Subclass Vetigastropoda
Overview
This large and diverse group is entirely marine, and includes keyhole and slit-limpets (Fissurellidae), abalone (Haliotidae), slit shells (Pleurotomariidae), and top shells (Trochidae). Many of these forms were previously included in "Archaeogastropoda" and this taxonomic term is still sometimes used in a narrow sense as a synonym for “vetigastropod” (e.g., Frýda et al., 2019). Many patellogastropods show numerous primitive features, such as paired ctenidia and shells with slits or other secondary openings. Unlike all other gastropods, the shell is nacreous and has an iridescent, mother-of-pearl sheen in many taxa. Coiled vetigastropods have a circular operculum, but this is absent in those taxa that have a limpet- or abalone-like shell shape. The operculum is thick and calcified in many members of the Trochoidea (families Turbinidae, Phasianellidae).
Species of Vetigastropoda have a paired or single ctenidium. Most living vetigastropods have small eggs that hatch into non-feeding (lecithotrophic) larvae, with protoconchs of one or fewer whorls.
It was once thought that the presence of apertural indentations, slits, or other openings on the shell (such as in true limpets and abalones) was indicative of paired ctenidia and signified “primitive” water circulation through the mantle cavity. This turned out to be misleading (as these shell features also occur in some extant caenogastropods that have only a single gill). While it is true that some living vetigastropods show the most primitive anatomical arrangement of any living gastropods, most living vetigastropods also share several derived anatomical characters.
Pleurotomariid-like forms occur in the Upper Cambrian, and trochoid-like forms in the Ordovician (Rohr and Frýda, 2001); some of these may represent stem-group vetigastropods. The first unambiguous vetigastropods probably occur in the Silurian. Vetigastropods are diverse and frequently well-preserved in Devonian and later Paleozoic rocks.
Examples of Vetigastropoda
Abalones: Family Haliotidae
By far the most famous of the Vetigastropoda are the abalones (family Haliotidae, genus Haliotis). They usually live on hard substrates in moderately shallow marine environments, and feed on macroalgae, seagrass, and diatoms.

Shell (dorsal and ventral views) of Haliotis rufescens from California (PRI 70105). Image by Jonathan R. Hendricks for the Digital Atlas of Ancient Life project.
These large snails, which have a series of holes in their shell, are edible and considered by some a delicacy. As shown in the video below, attempts are currently underway in some areas to farm abalone.
In addition to being targets of human exploitation for food in many parts of the world, the nacreous layer of the inner shell has long been a source of mother-of-pearl, which has been widely used as a material for jewelry, decoration, clothing buttons, and other uses.

Modern Haliotis irus from New Zealand (PRI 70101). The top layer of shell material has been removed from the right side of the specimen, revealing the nacreous layer beneath. Image by Jonathan R. Hendricks for the Digital Atlas of Ancient Life project.
Keyhole Limpets: Fissurellidae

Very large specimen of the modern keyhole limpet Megathura crenulata, collected at Mission Beach, San Diego, California (collection of the Paleontological Research Institution). Additional keyhole limpet specimens are shown in the background. Photograph by Jonathan R. Hendricks for the Digital Atlas of Ancient Life project.
Keyhole limpets (Fissurellidae), a group of Vetigastropoda, have a hole at the top of their cap-like shell. Their name comes from the shape of this hole.
Slit Snails: Pleurotomaria sp.
Subclass Neomphalina

Subclass Patellogastropoda

Shell of the modern limpet Lottia pelta, collected off Orcas Island, San Juan County, Washington (PRI 70356). Image by Jonathan R. Hendricks for the Digital Atlas of Ancient Life project.
Patellogastropoda includes the “true limpets,” which have low taxonomic diversity compared to other gastropod groups. These are the limpets that are commonly observed at rocky intertidal habits, where individuals cling to rocks as they are pounded by the surf.

A limpet (Testudinalia testudinalis) on a rock at low tide on Appledore Island, Maine. Photograph by Warren D. Allmon.
Nearly all living species are marine, and the group is widespread on rocky coasts, though not all are associated with shallow water. The shell is “limpet-shaped” and never coiled. Patellogastropods have the most diverse shell microstructure of any group of gastropods; the shell is never nacreous but otherwise may consist of a cross-lamellar, prismatic, or foliated structure. The protoconch is uncoiled and tubular, and larvae are lecithotrophic. An operculum is present in larvae, but is absent in juveniles and adults. There is either a single ctenidium, or none.
Patellogastropoda is among the gastropod groups previously referred to as “archaeogastropods.” They are now generally considered to be the sister group to all other living gastropods. They show many primitive characters, but their shell shape is likely derived from coiled ancestors. They may not be monophyletic. (The taxon Eogastropoda has been proposed to unite living patellogastropods with their extinct coiled ancestors.)
The limpet shape repeatedly evolved in all major gastropod groups (Vetigastropoda, Patellogastropoda, Neritimorpha, Heterobranchia including pulmonates, and Caenogastropoda). Despite their “basal” status among living gastropods, the oldest definite patellogastropods occur in the Triassic.
Content usage
Usage of text and images created for DEAL: Text on this page was written by Warren D. Allmon or Jonathan R. Hendricks. Original written content created by Warren D. Allmon or Jonathan R. Hendricks for the Digital Encyclopedia of Ancient Life that appears on this page is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Original images and diagrams created by Jonathan R. Hendricks or Warren D. Allmon are also licensed under Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
Content sourced from other websites: Attribution, source webpage, and licensing information or terms of use are indicated for images sourced from other websites in the figure caption below the relevant image. See original sources for further details. Attribution and source webpage are indicated for embedded videos. See original sources for terms of use. Reproduction of an image or video on this page does not imply endorsement by the author, creator, source website, publisher, and/or copyright holder.
Adapted images. Images that have been adapted or remixed for DEAL (e.g., labelled images, multipanel figures) are governed by the terms of the original image license(s) covering attribution, general reuse, and commercial reuse. DEAL places no further restrictions above or beyond those of the original creator(s) and/or copyright holder(s) on adapted images, although we ask that you credit DEAL if reusing an adapted image from the DEAL website. Please note that some DEAL figures may only be reused with permission of the creator(s) or copyright holder(s) of the original images. Consult the individual image credits for further details.