


Chapter contents:
Conservation Paleobiology
– 1. Foundations in Conservation Biology ←
– 2. Approaches in Conservation Paleobiology
–– 2.1 Near-time Approaches
–– 2.2 Deep-time Approaches
– 3. Deliverables from Conservation Paleobiology
– 4. Barriers to Applying Conservation Paleobiology
Above image by Ritiks; Creative Commons Attribution-Share Alike 3.0 Unported license.
Introduction
The field of conservation biology has deep roots in human philosophy but was only formalized as a field of scientific study in the 1980s. In his seminal paper—entitled “What is conservation biology?”—Michael Soulé (1985) detailed the postulates defining the field. Soulé put forth these postulates, which he considered as either functional or normative, to define the goals and values of the young field. More recently, conservation biologists have expanded the set of goals they address and many now include goals that benefit people alongside traditional goals geared strictly towards the benefit of nature. Conservation paleobiologists can provide important insights to address both the traditional and more expansive goals of conservation biology.
Soulé's functional postulates
Soulé’s four functional postulates and their corollaries describe the ecological and evolutionary mechanics underlying biodiversity. The postulates are: (1) Many of the species that constitute natural communities are the products of coevolutionary processes; (2) Many, if not all, ecological processes have thresholds below and above which they become discontinuous, chaotic, or suspended; (3) Genetic and demographic processes have thresholds below which nonadaptive, random forces begin to prevail over adaptive, deterministic forces within the populations; and, (4) Nature reserves are inherently disequilibrial for large, rare organisms. We will take a closer look at these functional postulates then turn our attention to the normative postulates.
The first postulate, that species in communities are the products of coevolution, is more readily understood with consideration of its four corollaries. These corollaries are: (1) species are interdependent; (2) many species are highly specialized; (3) extinction of keystone species can have long-range consequences; and, (4) introductions of generalists may reduce diversity. Put another way, this postulate and its corollaries could be summarized as: species do not exist in a vacuum; their past and present ecologies are important. The foundations of this postulate are also related to successional theory, which was a major focus of ecology in the first half of the twentieth century. In successional theory, communities themselves are considered to have some of the characteristics of individual organisms, as communities are considered to have “climax” stages, where they are essentially mature. The important concept is the deterministic nature of species inclusion in communities and their tight-knit functioning. As suggested by the corollaries and the postulate itself, removal of a single species from the community, or addition of one that was not there originally, threatens to throw the entire community into disarray. Though this may not be true of every species, it does become increasingly true for each species that is removed, as the resilience of the community will be depleted over time.
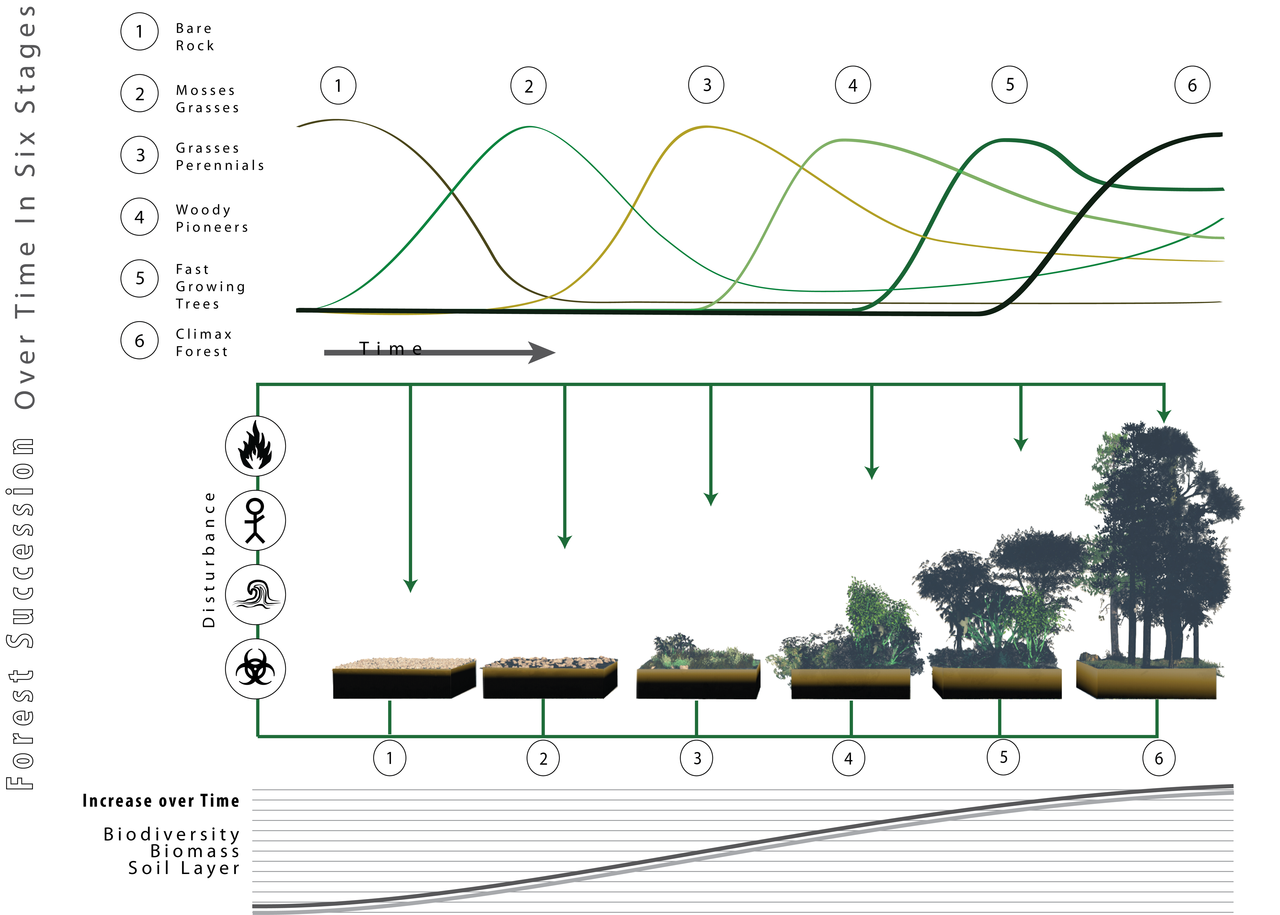
The succession of a forest ecosystem over time. The top panel shows the relative abundances of different groups of organisms as the forest develops. The middle panel gives a visual representation of the ecosystem at its different stages. The bottom panel shows the ever increasing biomass, biodiversity, and soil profile as the forest develops. As described by successional theory, and implied by Soulè's postulates, the composition of the climax community is strongly linked to the species present during the early stages of succession. Image by LucasMartinFrey; Creative Commons Attribution 3.0 Unported license.
{kind=link}
The second postulate, that ecological processes have threshold levels, acknowledges that these processes are often scale dependent. Relating again to succession, the first corollary to this postulate is that temporal continuity of habitats and successional stages depends on size. Generally, this can be interpreted to mean that if communities do not have enough space, as required for different stages of community development, they will not be able to function at capacity and the community will not reach its climax stage. Another way the successional process can break down, as described by the second corollary to this postulate, is with sudden increases to population sizes. For example, if the population of an herbivore (e.g., deer) grows rapidly—perhaps because humans have hunted all the large predators that kept the herbivore population in check—species consumed by the herbivore will be depleted rapidly. These sudden changes in population can ultimately lead to decreases in diversity in the community as one, or a few, species become dominant. There are both upper and lower limits for different ecological processes and when these limits are breached community dynamics change, and so do the communities themselves.

In this example, removal of the predator (i.e., wolves) allows the population of its prey (i.e., deer) to grow rapidly. The rapid increase in deer population results in the depletion of the plants they consume. Left unchecked, this process can dramatically alter ecosystems and their functioning. Image by Ccarroll17; Creative Commons Attribution-Share Alike 4.0 International license.
{kind=link}
The third postulate also deals with thresholds, this time at the level of genetics and demographics, and is largely focused on maintaining genetic diversity at high enough levels to avoid maladaptive consequences. Again, we can turn to the corollaries of this postulate for more details, though they can be summarized (over simplistically) by saying: bad things happen when populations get small. The first corollary defines small as populations under 50 or 100 individuals for outbreeding organisms (organisms where individuals exchange genetic material during the reproductive process). At or below these levels, genetic diversity drops, and the population will begin to exhibit genetic malformities and will lose its capacity to adapt to changing circumstances. As described in the second corollary, this loss of fitness (i.e., capacity to reproduce) is due to the process of genetic drift, or the random recombination of genetic information during reproduction. In large populations, genetic drift can produce useful variability in the population but in smaller populations, those effects become a detriment. These effects are summarized in the third corollary, which states that natural selection (learn more about Natural Selection) is less effective in populations under 50 or 100 individuals because of genetic drift and the collective loss of genetic variation. Taken in combination, the second and third postulates begin to define the scope of the effort required to maintain functioning communities.
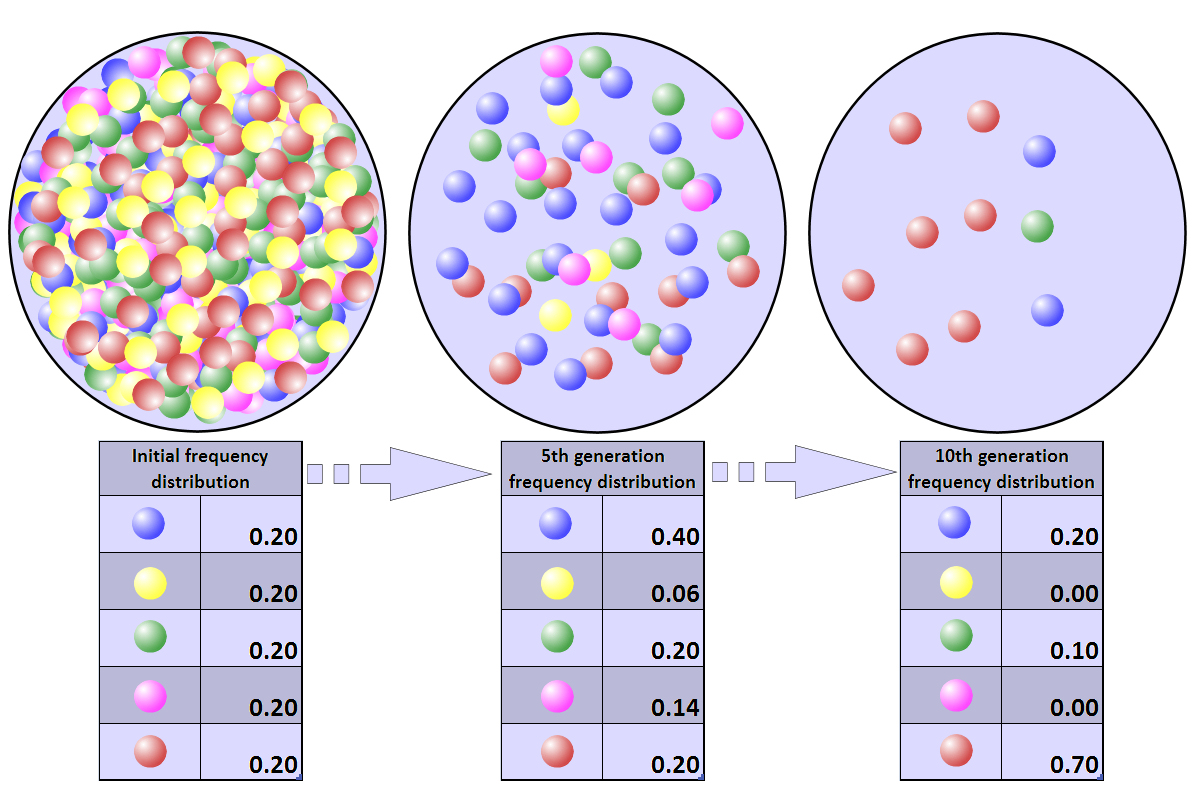
In this figure, the five colored balls represent genetic diversity. As the population gets smaller (moving from left to right), the genetic diversity in the population declines. As diversity declines, so too does the capacity of the population to respond to changing conditions. Image by Professor marginalia; Creative Commons Attribution-Share Alike 3.0 Unported license.
{kind=link}
The fourth postulate further defines this scope, stating that nature reserves do not represent an equilibrium state for large, rare organisms. Large organisms tend to have large ranges and their populations are sparser than smaller organisms. This means that to naturally maintain a population above 50 or 100 individuals, and for those individuals to interact naturally with other species in the community, very large tracts of land (or water) are required. Usually, the space available for a reserve is much smaller than would be natural. Consequently, large organisms in reserves are held at higher densities than in the wild, which affects their interactions with each other and other species in their community. Though it is not “natural,” such circumstances are still typically preferred to the alternative, which is to let the populations of large organisms decline unabated.
Taken together, the functional postulates described by Soulé have defined much of the conservation and restoration that has taken place in the last few decades.
Soulé's normative postulates
Underlying and complementing the functional postulates, the normative postulates provide us with the “why?” behind the course of action prescribed by Soulé. Whereas the functional postulates are based on the best available ecological knowledge, the normative postulates are much more philosophical. The four normative postulates are: (1) Diversity of organisms is good; (2) Ecological complexity is good; (3) Evolution is good; and, (4) Biotic diversity has intrinsic value. The first three postulates are very similar, and so we will consider them as a group before exploring the meaning of intrinsic value.
What does it mean for something to be good? Every person likely has a specific interpretation of “good” as it relates to their own personal experiences. Still, most definitions include desirability, utility, or value, in some sense. As applied to diversity, ecological complexity, and evolution, the connection between these concepts is not clear-cut. As described by Soulé, the goodness of these things comes from a deep visceral feeling—one that is shared by many biologists. It can also be helpful to define how these concepts are good by examining their opposites. Diversity—at all levels—provides variety and redundancy. It is this variety that allows natural selection to work and evolution to unfold. Redundancy means that systems can accommodate changes without collapsing. Without the variety and redundancy provided by diversity, most ecosystems would collapse. Much in the same way, ecological complexity leads to resilience of natural systems. It describes the interwoven, deep evolutionary connects between organisms. Without ecological complexity, ecosystems would merely be loose collections of species incapable of functioning as a cohesive unit. Evolution is the process by which species adapt and change. Without evolution, species would not be able to adapt to changing environmental conditions and would go extinct. Though much more can be said about each of these important concepts, by considering their absences we can begin to appreciate their importance.
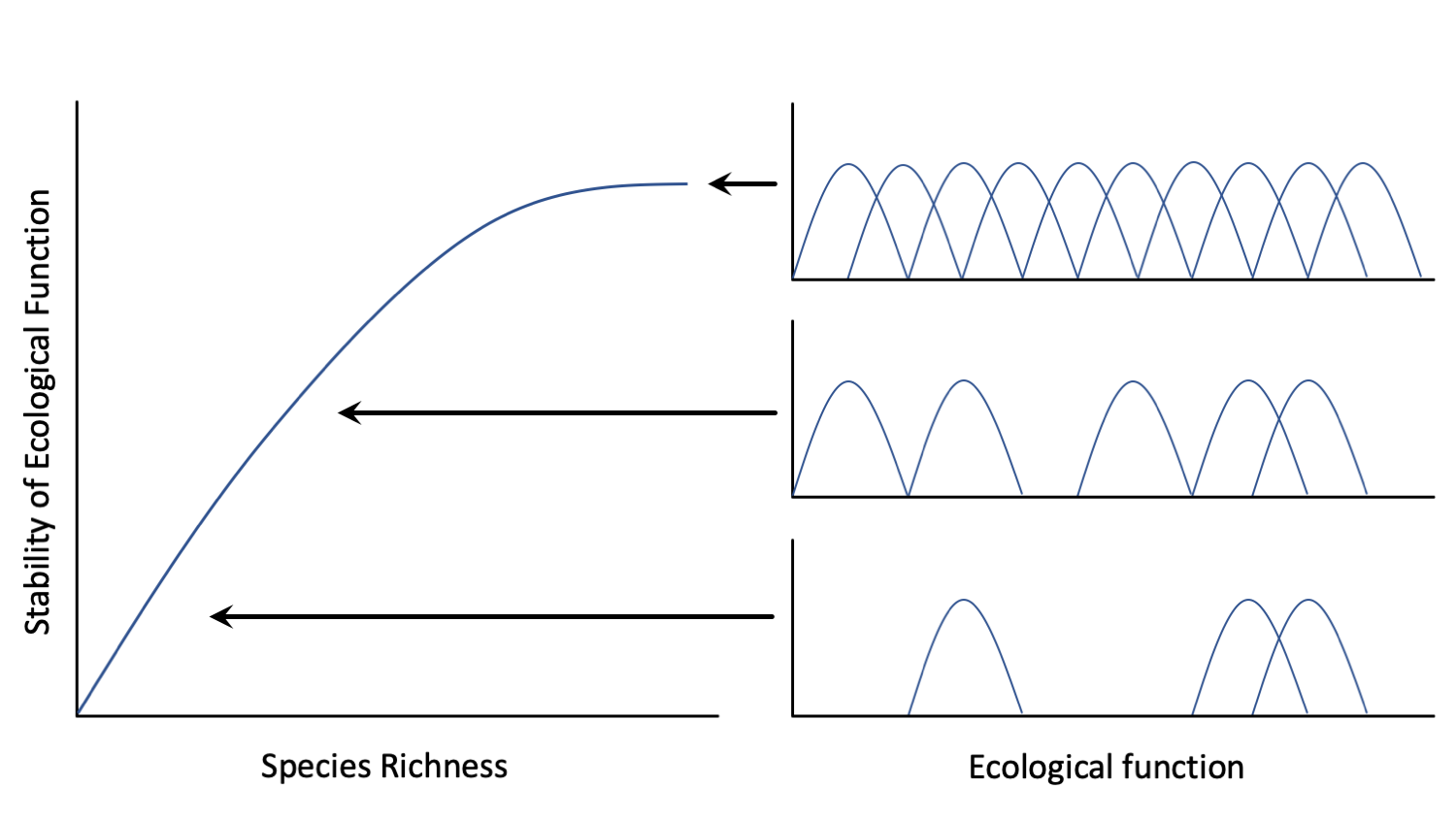
The stability of an ecosystem and its ability to function is often directly linked to the diversity of species occupying the ecosystem. When more species are present (as represented by the normal curves in the right side of the figure), there is often greater redundancy, and thus stability, in the functioning of the ecosystem. Redrawn from Peterson et al. (1998) in Ecosystems, after the concept by Ehrlich and Ehrlich (1981).
Biotic diversity (or biodiversity) has intrinsic value. As you may have recognized in the preceding discussion, this fourth postulate is implied by each of the first three normative postulates. Removal of diversity, loss of ecological complexity, and the cessation of evolution all ultimately lead to the depletion of biodiversity. Why is that a bad thing? As we will see in the next section, the loss of biodiversity has important implications for people, but, first and foremost, the loss of biodiversity is bad because species—and by extension, all individuals—have intrinsic value. Intrinsic value is the innate value conferred simply through existence. That is, every species is good simply because it exists, and has a right to exist, without a need for an external value. This is a challenging concept to digest and again we can consider a contrast to provide clarity. The opposite of intrinsic value is extrinsic value. Extrinsic value is externally defined, by people, based on the usefulness of, or economic market for, a species and/or its byproducts. In the theoretical realm of conservation biology, existence merits conservation on the basis of intrinsic value alone, regardless of any and all extrinsic valuation. By extension of these postulates, and as a consequence of them, the long-standing goal of conservation biology has been to maintain and restore natural ecosystems, and the species in them (i.e., historical fidelity).
The expanding set of goals in conservation biology
Scientific fields change over time and conservation biology is no different. As discussed above, the primary goal of conservation biology has been to maintain and restore natural ecosystems and the species they hold. Many in conservation biology adhere to this goal and have been reluctant to include extrinsically defined (i.e., human-facing) goals. Rather than include humans and their actions as a part of nature, those pursuing the goal of historical fidelity in natural ecosystems exclude humans from the equation. In the United States, the National Park system is a good example of this approach: humans are allowed to visit but the lands remain undeveloped. On top of this, conservation and restoration efforts consciously attempt to maintain these complex systems in the same state they have existed historically. Recognizing that this will not always be possible and, because they believe people should be factored into the plan, others have started working towards a more expansive set of goals.

Yellowstone National Park, the archetypal nature reserve adhering to the conservation principle of historical fidelity. Image by Ed Austin/Herb Jones (public domain).
{kind=link}
One of the major criticisms of traditional conservation is that is adheres to an unnecessarily strict conception of how an ecosystem should exist. In many cases, “natural” is synonymous with the way the ecosystem was first encountered by non-native explorers. That is, “natural” is set to an arbitrary time in the past, and from what is often a biased perspective (i.e., that of the non-native explorer). Though there can be merit to this approach, it also does not permit flexibility during times of environmental change, such as the episode of climate change we are currently experiencing. Learning from the geohistorical record, we know that species either adapt to the new conditions, move to new places to track the environmental conditions that they prefer, or die. By rigidly attempting to maintain the same ecosystem and the same set of species in a given location, conservation biologists may be setting themselves up to fail, despite the best of intentions.
Fortunately, many conservation biologists have recognized this threat and have adopted a broader set of goals. Some of these goals include human interests, but many adhere to the normative postulates detailed by Soulé in 1985. Those including human interests tend to focus on the concept of ecosystem services, which are the benefits people derive from ecosystems. These are the means by which the extrinsic value of ecosystems tends to be assessed.

Ecosystem services, the benefits derived by people from the natural world, and their management are influenced by many factors, including the benefit provided, the resources required to attain that benefit, and the drivers of change affecting it (left). At right, we can see a handful of these factors influencing the service of food, fuel, and materials provisioning. Images from the United States Environmental Protection Agency - EnviroAtlas (public domain).
Ecosystem services come in four varieties: provisioning, regulating, supporting, and cultural. Provisioning services include things like production of food and potable water. Regulating services are often processes like climate cycling and pollination. Supporting services include nutrient (e.g., nitrogen, phosphorous, carbon) cycling and production of oxygen. Cultural services tend to be more spiritual or recreational and deal with how humans experience nature. In the expanded set of conservation goals, maintaining and maximizing ecosystem services are prominent. Though ecosystem services are often discussed as a single entity and in relatively abstract terms, conservation and restoration efforts in local areas often focus on specific services that are desirable to stakeholders in the area. For example, on the coasts, where sea level rise and increasing storm severity threaten the shoreline, the regulating services of coastal protection or shoreline stability—imparted by coral reef and mangrove ecosystems, respectively—may be the focus of conservation actions. In many cases, however, the maintenance of ecosystem services is thought about more diffusely, so as to retain the option value of the services for the future.
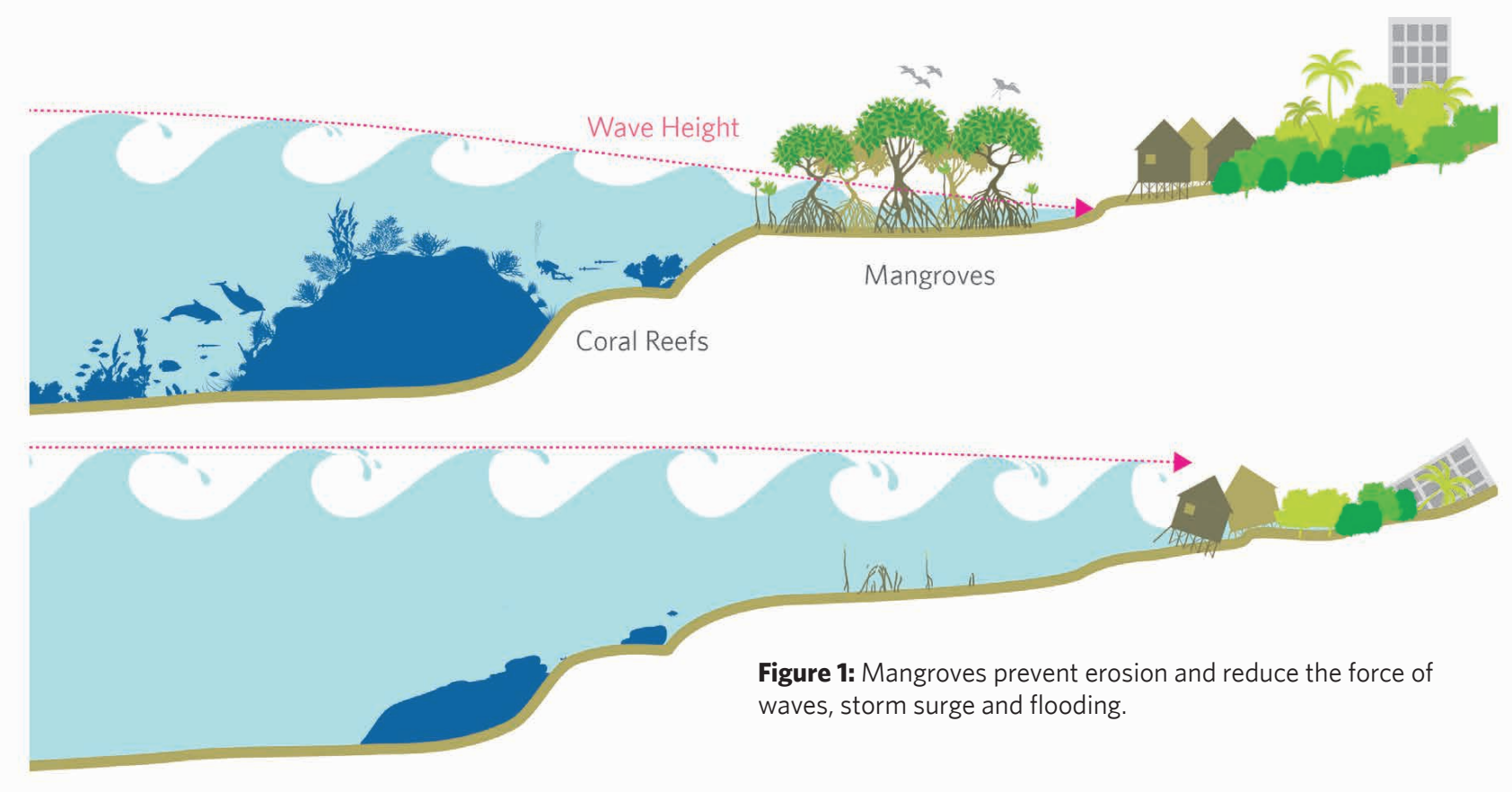
When natural shoreline ecosystems are present, such as mangrove and coral reef systems, they protect and stabilize the shoreline by dissipating wave energy (top panel). Without these systems in place, waves directly contact the shoreline and can be destructive (bottom panel). Figure from Losada et al. (2018).
Still, not all of the more expansive set of goals include human interests. For example, in place of maintaining the exact same ecosystem, efforts instead can be spent towards maximizing the biodiversity in an ecosystem or on mimicking the structure and/or function of the historical ecosystem. These two goals, in particular, reflect the normative postulates presented by Soulé. Maximizing biodiversity matches well with the postulates stating that diversity of organisms is good and biotic diversity has intrinsic value. Mimicking structure and/or function is in acknowledgment of the statement that ecological complexity is good. From a normative perspective, the expanded set of goals in conservation biology is not necessarily any different from the traditional goal.
On a functional level, there is more contention between the traditional and newer perspectives in conservation biology. As outlined by Soulé, ecosystems are a reflection of their history and species very often have evolved in concert with the other species in their ecosystem. With species geographic distributions shifting and the climate changing, novel ecosystems—ecosystems composed of sets of species that have not historically occurred together—are becoming increasingly more common. In these novel ecosystems, it remains to be seen whether they can be structured and perform functions in the same way as historical ecosystems. Understanding how novel ecosystems compare to historical ecosystems is now a pressing issue in conservation biology.

In conservation practice, the goals of both traditional and new conservation can be addressed. Though they are often treated as two points of a dichotonomy, these perspectives and their sets of goals are more truly end members of a spectrum of human- and nature-oriented goals. Figure by Jansen Smith.
Though the traditional and expanded set of goals in conservation seem to be at odds, it is important to recognize that they represent points on a spectrum and not two parts of a dichotomy. Ultimately, all conservation biologists desire that for which they are named: they want biology to be conserved. The methods applied and the motivating goals may differ, but all are working towards the same thing. These differences have caused strife, and will continue to; however, the issues facing the world are complex and a one-size-fits-all approach will fail. To add to the Soulé’s postulates, we might also consider: a diversity of approaches to conservation biology is good.
Putting the Paleo into Conservation
Conservation paleobiology can help to diversify the approaches to conservation by offering more data, longer timescales, and different perspectives. Whether applied to the traditional or expanded set of goals, geohistorical records provide invaluable context for the world around us today.
Soulé placed a heavy emphasis on the evolutionary history of organisms in ecosystems, noting that organisms cooccurring in ecosystems likely evolved together and that their interactions define many aspects of the ecosystem. Geohistorical records can be used to further explore these evolutionary relationships, as the fossil record can show us how the species interacted and evolved over the preceding thousands and millions of years. Alternatively, the fossil record can be studied to examine the assumption that evolutionary history matters. As mentioned above, the geohistorical record, and more specifically the fossil record, demonstrates that ecosystems are constantly changing. By examining the ways communities form and break apart in the fossil record, we can better understand the importance of community structure and dynamics in modern ecosystems.

A subset of communities from different ecosystems found in the geohistorical record, including Devonian snails, crinoids, and brachiopods (leftmost), Cretaceous plants and insects (left-central), Miocene mammals (right-central), and Quaternary snails and clams (rightmost). Leftmost and rightmost images by Jansen Smith. Left-central image from Labandeira et al. (2002) in the Proceedings of the National Academy of Sciences under the PNAS License to Publish (Copyright (2002) National Academy of Sciences). Right-central image by Ammodramus; Creative Commons CC0 1.0 Universal Public Domain Dedication.
{kind=link}
The utilization of geohistorical records can also help define what is meant by the traditional goal of maintaining the same ecosystem. Without records from the past, the target for the “same ecosystem” may not accurately reflect the dynamism of an ecosystem over time. Instead of choosing the conditions in an ecosystem from an arbitrary point in time (e.g., prior to European settlement), geohistorical records can be used to assess the degree to which an ecosystem and its community were stable over time and, fundamentally, which species were present. For example, in the Galápagos, pollen grains preserved in bog sediments have been used to determine whether populations of six plant species are native to the islands and should be maintained (read more in the section on Conservation Paleobiology). Though records are not be available in all places and are subject to taphonomy and time-averaging, the value provided by geohistorical records remains high when they are available and applicable.
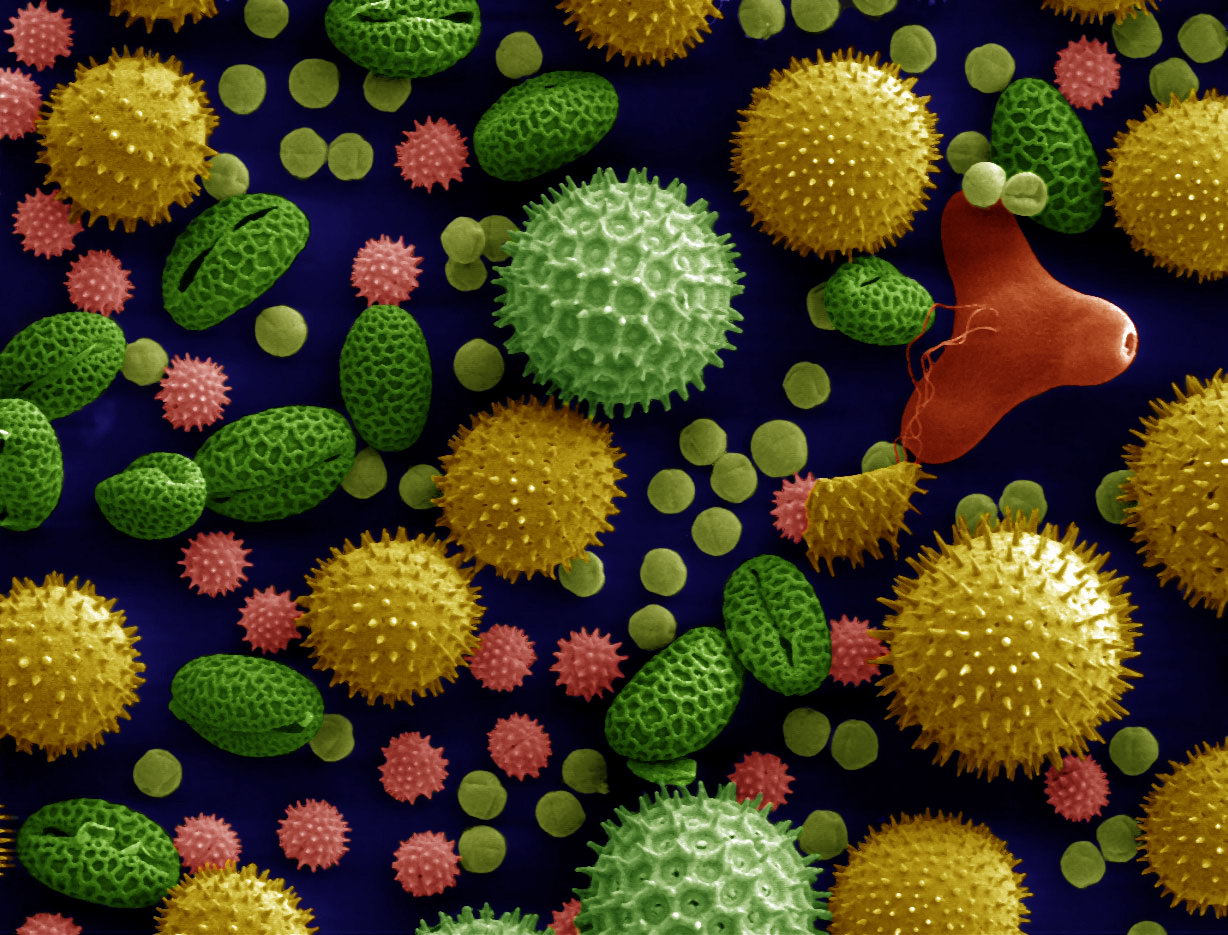
Colorized pollen grains viewed under a scanning electron microscope. Fossilized pollen, and other geohistorical records, can contribute invaluable information to the decision-making process in conservation. Image by Dartmouth Electron Microscope Facility, Dartmouth College (public domain).
{kind=link}
Much as conservation paleobiology can contribute to achieving the goal of historical fidelity, so too can it address the more expansive set of goals in conservation biology today. For instance, if the goal is to maximize biodiversity, the geohistorical record of biodiversity can be used to set standards for conservation and restoration practice. Similarly, if the goal is to mimic historical structure and function, the geohistorical record can be used—much as it is when the goal is historical fidelity—to add a longer temporal perspective that is more inclusive of the variability inherent to every ecosystem. Most recently, conservation paleobiologists have begun to analyze the relationship between the traits organisms possess and the ecosystem services that tend to be imparted as a function of those traits. With these new trait-based approaches, the geohistorical record can also be applied to the newer conservation goals concerned with the maintenance and maximization of ecosystem services. Geohistorical records can be applied to most conservation goals, adding a valuable long-term perspective to conservation biology.
Continue to the next section to learn more about the specific approaches used by conservation paleobiologists to address conservation goals!
Concept check: See what you know!
What are Soulé's functional postulates?
(1) Many of the species that constitute natural communities are the products of coevolutionary processes; (2) Many, if not all, ecological processes have thresholds below and above which they become discontinuous, chaotic, or suspended; (3) Genetic and demographic processes have thresholds below which nonadaptive, random forces begin to prevail over adaptive, deterministic forces within the populations; and, (4) Nature reserves are inherently disequilibrial for large, rare organisms
What are Soulè's normative postulates?
(1) Diversity of organisms is good; (2) Ecological complexity is good; (3) Evolution is good; and, (4) Biotic diversity has intrinsic value
What is an ecosystem service?
An ecosystem service is any benefit that humans acquire from the natural world, from drinking water to the simple pleasure of walking through a forest.
How are geohistorical records relevant to the traditional goals of conservation?
In traditional conservation, the goal is historical fidelity (or "naturalness"). Geohistorical records are often the only way to even know what "natural" truly is, in the sense that these records can predate human effects on ecosystems.
True or False: There is no way to reconcile the human- and nature-oriented goals of traditional and new conservation.
In the opinion of the author (Jansen Smith), this is false. The goals set in conservation should be situation dependent and can almost always accommodate both perspectives. What do you think?
References and Further Reading
Barnosky, A. D., E. A. Hadly, P. Gonzalez, J. Head, P. D. Polly, A. M. Lawing, et al. 2017. Merging paleobiology with conservation biology to guide the future of terrestrial ecosystems. Science, 355: eaah4787.
Ehrlich, P., and A. Ehrlich. 1981. Extinction: the causes and consequences of the disappearance of species. New York, Random House. 305 p.
Groom, M. J., G. K. Meffe, C. R. Carroll, and S. J. Andelman. 2006. Principles of conservation biology. Sunderland: Sinauer Associates. 779 p.
Losada, I. J., P. Menéndez, A. Espejo, S. Torres, P. Díaz-Simal, S. Abad, M. W. Beck , S. Narayan, D. Trespalacios, K. Pfiegner, P. Mucke, L. Kirch. 2018. The global value of mangroves for risk reduction. Technical Report. Berlin, The Nature Conservancy. 44 p.
Marris, E. 2011. Rambunctious garden: saving nature in a post-wild world. New York, Bloomsbury Publishing. 224 p.
Peterson, G., C. R. Allen, and C. S. Holling. 1998. Ecological resilience, biodiversity, and scale. Ecosystems, 1: 6-18.
Soulé, M. E. 1985. What is conservation biology? BioScience, 35: 727-734.