


Section by
Elizabeth J. Hermsen, Paleontological Research Institution, Ithaca, New York.
Section contents
- Paleoclimate estimation with plant fossils ←
Related pages & background reading
Topics covered on this page
Introduction
While many paleobotanists study plant structure and evolution, some are more interested in using plants to study paleoecology and paleoclimate. Paleobotanists have come up with ways to use plants or the features of plants to estimate aspects of past climate like mean annual temperature (average yearly temperature) and mean annual precipitation (average yearly rainfall and snowfall). Thus, plants play an important role in telling us about the environment of the past and in helping us to predict what the environment of the future might be like as the Earth warms. So, how do paleobotanists use plants to study temperature are rainfall in the past? There are several approaches, which are summarized below.
Abbreviations
Below is a table abbreviations used on this page for quick reference.
| Abbreviation | Meaning |
|---|---|
| CA | coexistence approach, a nearest living relative (NLR) method of estimate paleoclimate |
| CLAMP | Climate Leaf Analysis Multivariate Program, a multivariate method of estimating paleoclimate using leaf physiognomy |
| DiLP | digital leaf physiognomy, a multivariate method of estimating paleoclimate using leaf physiognomy |
| LAA | leaf area analysis, a univariate method for estimating mean annual precipitation (MAP) using leaf physiognomy |
| LMA | leaf margin analysis, a univariate method for estimating mean annual temperature (MAT) using leaf physiognomy |
| MAT | mean annual temperature (average yearly temperature) |
| MAP | mean annual precipitation (average yearly precipitation, like rainfall and snowfall) |
| NLR | nearest living relative |
Nearest living relative (NLR) methods
Nearest living relative methods (NLR) rely on the principle of physiological uniformitarianism, basically the idea that closely related taxa have maintained similar environmental tolerances and requirements through time. Thus, the nearest living relatives of fossil plants can provide information on past climates. NLR methods require identification of taxa in order to estimate paleoclimatic parameters. One benefit of NLR methods over leaf physiognomy (described below) is that NLR methods can be applied to a broader range of plant taxa and plant organ types; for example, NLR can be used on pollen and spore floras, fruit and seed floras, or wood, in addition to leaves.
One method of using NLR is to estimate paleoenvironmental parameters by looking for one or more indicator taxa in a paleobiota. For example, a common way to estimate minimum cold temperatures in an environment is to look for indicator taxa that have little capacity to survive in freezing conditions for an extended period of time, such as cycads, palms, or crocodiles.

Fossils of cold-intolerant organisms. Left: Blade of a palm leaf (Sabalites powellii, Eocene, Green River Formation, Wyoming, USA). Right: A juvenile alligator (Alligator, Pliocene, Gray Fossil Site, Tennessee, USA). Credit: Photos by E.J. Hermsen (DEAL).
A more systematic approach to using NLR is the coexistence approach (CA). This technique uses the overlap in climate tolerances of the nearest living relatives of plant taxa that occur in the same paleoflora to estimate the environmental parameters (for example, average yearly temperature and rainfall) of that paleoflora. Often, CA analyses are performed at the genus level.
NLR methods have potential drawbacks. The major potential problem with these methods is, of course, that the assumption of physiological uniformitarianism could be wrong for one or more taxa in an NLR analysis. Other critiques are connected to the specific methods used.
Leaf physiognomy
Leaf physiognomic methods are methods that rely on the physical characteristics of leaves in order to estimate paleoclimatic parameters such as mean annual temperature (MAT) and mean annual precipitation (MAP). One major advantage of leaf physiognomic methods is that they do not rely on correct plant identification in order to work. Instead, leaf physiognomic methods can be applied using leaf morphotypes, or groups of morphologically similar leaves that are not formally classified (discussed further below).
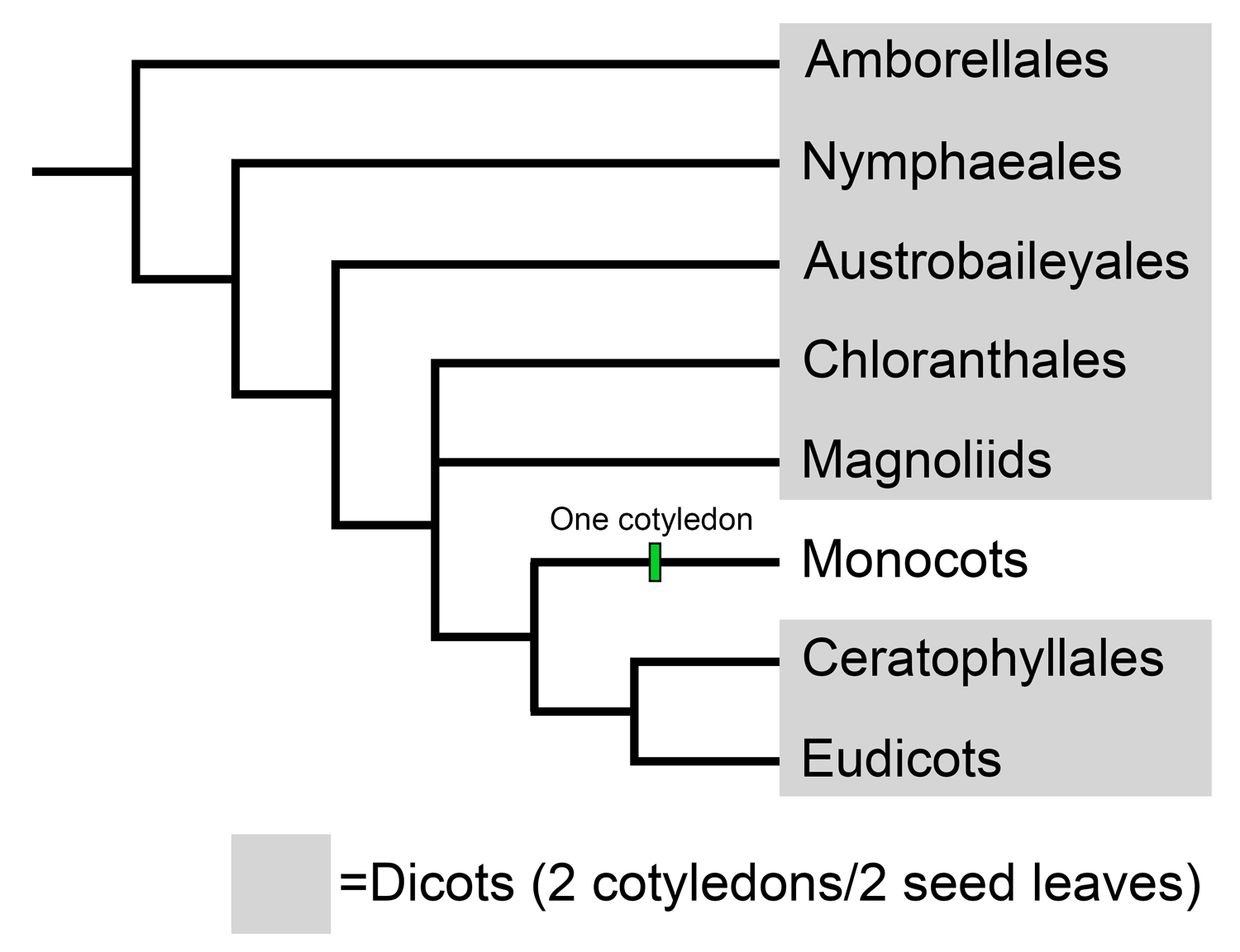
Leaf physiognomic methods have been developed for woody angiosperms (flowering plants) that are commonly called dicots. A dicot is an angiosperm that has an embryo with two cotyledons, or seed leaves. In modern angiosperm classification systems, the dicots are a paraphyletic group. Most dicots are eudicots, the largest group of angiosperms; however, some are in other groups, most notably the Magnoliids. Dicots tend to have leaves with reticulate venation. Leaf physiognomic methods exclude the leaves of herbaceous plants (non-woody plants) and monocots.

Reticulate leaf venation. Reticulate leaf venation in witch hazel (Hamamelis), a eudicot. Reticulate venation is venation that forms a network. Credit: E.J. Hermsen (DEAL).
Leaf physiognomic methods use formulas based on studies of modern floras in order to quantify the relationships between leaf traits and climate variables like temperature and precipitation. These formulas are then applied to fossil floras. There are two basic types of formulas:
- Univariate approaches use correlations between a single leaf trait—for example, leaf margin type or leaf area—and a single climatic variable such as temperature or precipitation. Common univariate methods include leaf margin analysis (LMA) for temperature and leaf area analysis (LAA) for precipitation.
- Multivariate approaches use several leaf traits simultaneously to estimate one or more climatic variables. The most commonly used multivariate method is Climate Leaf Analysis Multivariate Program (CLAMP). Digital leaf physiognomy (DiLP) is a more recently developed method now being applied to fossil floras.
For leaf physiognomic studies, ideally a representative sample of all leaves for a given locality (or even a given horizon at a given locality) should be taken. Nevertheless, leaf physiognomic analyses have been performed on historical collections or collections that have been made for systematic studies.

Collecting leaf fossils. Left: A quarry where leaf fossils have been collected. Right: Fossil leaves wrapped in toilet paper and ready for labelling. Credit: E.J. Hermsen (DEAL).
Leaf morphotypes
Angiosperm leaf floras (and other types of fossil floras) may be described based on taxonomic concepts, morphotype concepts, or a mixture of the two. A leaf morphotype is a group of fossil leaves that is united by a shared type of morphology (form) and venation pattern. Morphotypes are usually defined on a study-by-study basis using numbers or a combination of letters and numbers; in other words, there is no shared list of morphotypes that all paleobotanists use. A given morphotype may or may not correspond to a formally named species.

Morphotypes. The images above are examples of morphotypes from the Eocene Florissant flora (Colorado, U.S.A.). Left: Three examples of morphotype 29, a toothed leaf also assigned to the species Ulmus tenuinervis (a fossil elm). Right: Two examples of a leaf with an entire (smooth) margin assigned to morphotype 15, but with no formal classification. Credit: Reproduced from Allen et al. (2020) PaleorXiv (CC BY 4.0). Images modified from originals.
The idea behind morphotyping the leaves in a flora is to describe and categorize the different types of leaves present without necessarily trying to determine their systematic affinities (in other words, their relationships or formal classification). One of the major advantages of morphotyping is that it allows for the diversity within a leaf flora to be described and quantified whether or not the systematic affinities of every morphotype can be determined.
Morphotyping may be done using leaves from a single locality or from multiple localities (quarries or sites) that are considered to be part of the same fossil flora. Typically, if leaves come from more than one locality, they were all collected from the same rock unit. The steps in morphotyping a flora are as follows (adapted from the 1999 Manual of Leaf Architecture):
- Divide leaves into groups based on similarity. Focus first on the most reliable and easily observed characters of the leaf margin (untoothed or toothed margins, unlobed or lobed margins) and venation pattern (type of primary and secondary venation). After that, finer details can be examined to further differentiate among morphotypes. For more about angiosperm leaf architecture, see the Angiosperm leaf architecture page.
- Choose a specimen to serve as the reference specimen (also called an exemplar or holomorphotype) for the characteristics of the morphotype. The reference specimen should be well preserved and show the characteristic features of the morphotype.
- Assign each morphotype a number and describe it based on the reference specimen and other specimens. (A leaf morphotype template and example descriptions are provided in the 2009 edition of the Manual of Leaf Architecture.) If a morphotype has the characters of a named species, it can also be assigned to that species (in other words, the morphotype number and species name can be used in together).

Angiosperm leaf morphotypes. Portion of a multipanel figure showing examples of leaf morphotypes from the Eocene Florissant Formation flora (Colorado, U.S.A.). Credit: Reproduced from Allen et al. (2020) PaleorXiv, fig. 3 (CC BY 4.0). Image modified from original.

Variation in white mulberry leaves. Herbarium specimens of white mulberry (Morus alba), a species that has simple leaves with toothed margins that vary from unlobed to five-lobed. Left: Branch with leaves that have teeth but lack lobes. Right: Branch with leaves that have teeth and lobes. Credits: Morus alba, NY01527673 (New York Botanical Garden, via GBIF, CC BY 4.0); Morus alba, NY01527546 (New York Botanical Garden, via GBIF, CC BY 4.0).

Shade and sun leaves of white oak. The amount of light exposure a leaf receives during development can affect its final form. Examples of shade (left) and sun (right) leaves of white oak (Quercus alba). Note that the sun leaf has more lobes and deeper sinuses (valleys between lobes) than the shade leaf. Credit: Quercus alba (Steven J. Baskauf, via Bioimages, CC BY 4.0).
References & further reading
Note: Free full text is made available by the publisher for items marked with a green asterisk.
Academic articles & book chapters
* Allen, S.E., A.J. Lowe, D.J. Peppe, and H.W. Meyer. 2020. Paleoclimate and paleoecology of the uppermost Eocene Florissant flora (Central Colorado, USA), ver. 2. PaleorXiv. [Preprint] https://doi.org/10.31233/osf.io/xpm26
Allen, S.E., A.J. Lowe, D.J. Peppe, and H.W. Meyer. 2020. Paleoclimate and paleoecology of the latest Eocene Florissant flora (Central Colorado, USA). Palaeogeography, Palaeoclimatology, Palaeoecology. https://doi.org/10.1016/j.palaeo.2020.109678
Eckenwalder, J.E. 1980. Foliar heteromorphism in Populus (Salicaceae), a source of confusion in the taxonomy of Tertiary leaf remains. Systematic Botany 5: 366–383. https://doi.org/10.2307/2418518
* Kidner, C.A., and S. Umbreen. 2010. Why is leaf shape so variable? International Journal of Plant Devleopmental Biology 4: 64–75. http://www.globalsciencebooks.info/Online/GSBOnline/images/2010/IJPDB_4(SI1)/IJPDB_4(SI1)64-75o.pdf
* Liu, Y., X. Li, G. Chen, M. Li, M. Liu, D. Liu. 2015. Epidermal micromorphology and mesophyll structure of Populus euphratica heteromorphic leaves at different development stages. PLoS ONE 10(9): e0137701. https://doi.org/10.1371/journal.pone.0137701
Mosbrugger, V., and T. Utescher. 1997. The coexistence approach—a method for quantitative reconstructions of Tertiary terrestrial palaeoclimate data using plant fossils. Palaeogeography, Palaeoclimatology, Palaeoecology 134: 61–86. https://doi.org/10.1016/S0031-0182(96)00154-X
* Nakayama, H., N.R. Sinha, and S. Kimura. 2017. How do plants and phytohormones accomplish heterophylly, leaf phenotypic plasticity, in response to environmental cues. Frontiers in Plant Science 8: 1717. https://doi.org/10.3389/fpls.2017/01717
Peppe, D.J., A. Baumgartner, A. Flynn, and B. Blonder. 2018. Reconstructing paleoclimate and paleoecology using fossil leaves. Pp. 289–317. In: D.A. Croft, D.F. Su, and S.W. Simpson, eds. Methods in Paleoecology: Reconstructing Cenozoic Terrestrial Environments. Springer Nature Switzerland AG, Switzerland. https://doi.org/10.1007/978-3-319-94265-0_13
* Peppe, D.J., D.L. Royer, B. Cariglino, S.Y. Oliver, S. Newman, E. Leight, G. Enikolopov, M. Fernandez-Burgos, F. Herrera, J.M. Adams, E. Correa, E.D. Currano, J.M. Erickson, L.F. Hinojosa, J.W. Hoganson, A. Iglesias, C.A. Jaramillo, K.R. Johnson, G.J. Jordan, N.J.B. Kraft, E.C. Lovelock, C.H. Lusk, Ü. Niinemets, J. Peñuelas, G. Rapson, S.L. Wing, I.J. Wright. 2011. Sensitivity of leaf size and shape to climate: global patterns and paleoclimatic applications. New Phytologist 190: 724–739. https://doi.org/10.1111/j.1469-8137.2010.03615.x
* Tiffney, B. 2008. Phylogeography, fossils, and Northern Hemisphere biogeography: The role of physiological uniformitarianism. Annals of the Missouri Botanical Garden 95: 135–143. https://doi.org/10.3417/2006199. Read/download free on Biodiversity Heritage Library: https://www.biodiversitylibrary.org/bibliography/702#/summary
Wilf, P. When are leaves good thermometers? A new case for Leaf Margin Analysis. Paleobiology 23: 373–390. https://doi.org/10.1017/S0094837300019746
Wing, S.L., and D.R.Greenwood. 1993. Fossils and fossil climate: The case for equable continental interiors in the Eocene. Philosophical Transactions: Biological Sciences 341: 243–252. https://doi.org/10.1098/rstb.1993.0109
* Wolfe, J.A. 1979. Temperature parameters of humid to mesic forests of eastern Asia and relation to forests of other regions of the Northern Hemisphere and Australasia. United States Geological Survey (USGS) Professional Paper 1106, 37 pgs. https://doi.org/10.3133/pp1106
* Wolfe, J.A. 1993. A method of obtaining climatic parameters from leaf assemblages. United States Geological Survey (USGS) Bulletin 2040, 71 pgs. https://doi.org/10.3133/b2040
Wolfe, J.A. 1995. Paleoclimate estimates from Tertiary leaf assemblages. Annual Review of Earth and Planetary Science 23: 119–142. https://doi.org/10.1146/annurev.ea.23.050195.001003
Books, textbooks & manuals
* CLAMP online: Climate Leaf Analysis Multivariate Program. http://clamp.ibcas.ac.cn/CLAMP_Home.html
* Ellis, B., D.C. Daley, L.J. Hickey, K.R. Johnson, J.D. Mitchell, P. Wilf, and S.L. Wing. 2009. Manual of Leaf Architecture. Comstock Publishing Associates, Cornell University Press, Ithaca, New York. Free PDF on ResearchGate (shared with Creative Commons Attribution-NonCommercial license, CC BY-NC 4.0): https://www.researchgate.net/publication/270216727_Manual_of_Leaf_Architecture
* Leaf Architecture Working Group (A. Ash, B. Ellis, L.J. Hickey, K. Johnson, P. Wilf, and S. Wing). 1999. Manual of Leaf Architecture: Morphological description and categorization of dicotyledonous and net-veined monocotyledonous angiosperms. Smithsonian Institution, Washington, DC. Free PDF from P. Wilf, Pennsylvania State University: https://personal.ems.psu.edu/~pdw3/1999_MLA.pdf
Websites
* Burke Museum Paleobotany Project: Morphotyping. https://www.burkemuseum.org/static/Republic/background6579.html
* Wagner, S. 2002. Castle Rock fossil rainforest: Identification of fossil leaves using morphotype process. Paleocurrents.com. http://www.paleocurrents.com/castle_rock/docs/morphotype_id_process.html
* Wagner, S. 2002. Castle Rock fossil rainforest: Challenges in morphotyping. Paleocurrents.com. http://www.paleocurrents.com/castle_rock/docs/morphotype_challenges.html
Content usage
Usage of text and images created for DEAL: Text on this page was written by Elizabeth J. Hermsen. Original written content created by E.J. Hermsen for the Digital Encyclopedia of Ancient Life that appears on this page is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Original images created by E.J. Hermsen are also licensed under Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
Content sourced from other websites: Attribution, source webpage, and licensing information or terms of use are indicated for images sourced from other websites in the figure caption below the relevant image. See original sources for further details. Attribution and source webpage are indicated for embedded videos. See original sources for terms of use. Reproduction of an image or video on this page does not imply endorsement by the author, creator, source website, publisher, and/or copyright holder.
Adapted images. Images that have been adapted or remixed for DEAL (e.g., labelled images, multipanel figures) are governed by the terms of the original image license(s) covering attribution, general reuse, and commercial reuse. DEAL places no further restrictions above or beyond those of the original creator(s) and/or copyright holder(s) on adapted images, although we ask that you credit DEAL if reusing an adapted image from the DEAL website. Please note that some DEAL figures may only be reused with permission of the creator(s) or copyright holder(s) of the original images. Consult the individual image credits for further details.
First officially released 28 September 2021; last updated 23 June 2022.