


Chapter contents:
Echinodermata
–– 1. Exclusively Fossil Taxa
–– 2. Crinoidea
–– 3. Asteroidea
–– 4. Ophiuroidea
–– 5. Echinoidea ←
–– 6. Holothuroidea
You can find 3D models of Echinoidea here!
This page was written by Jansen Smith and Jaleigh Q. Pier. It was last updated on May 27, 2020.
Above image: Photograph of live sea urchins in a touch tank. Image by Jonathan R. Hendricks, licensed under a Creative Commons Attribution-Share Alike 4.0 International License.
Class Echinoidea Snapshot
- Examples: sea urchins, sand dollars, sea biscuits, heart urchins
- Ecology: marine, mobile carnivores, herbivores, deposit feeders
- Key features of group: calcitic test, aristotle’s lantern, spines
- Diversity: ~4,221 living sp., ~10,375 extinct sp.
- Fossil record: Ordovician to Recent
Overview
Echinoidea comes from the Greek word ‘echinos’ meaning “spiny” and includes the sea urchins, sand dollars, and heart urchins. Echinoids are the sister taxa to holothuroidians within the Echinozoa clade. Like other extant echinoderms, species in this mobile class primarily exhibit pentaradial symmetry—some have evolved bilateral symmetry—and use a water vascular system for movement and food capture.
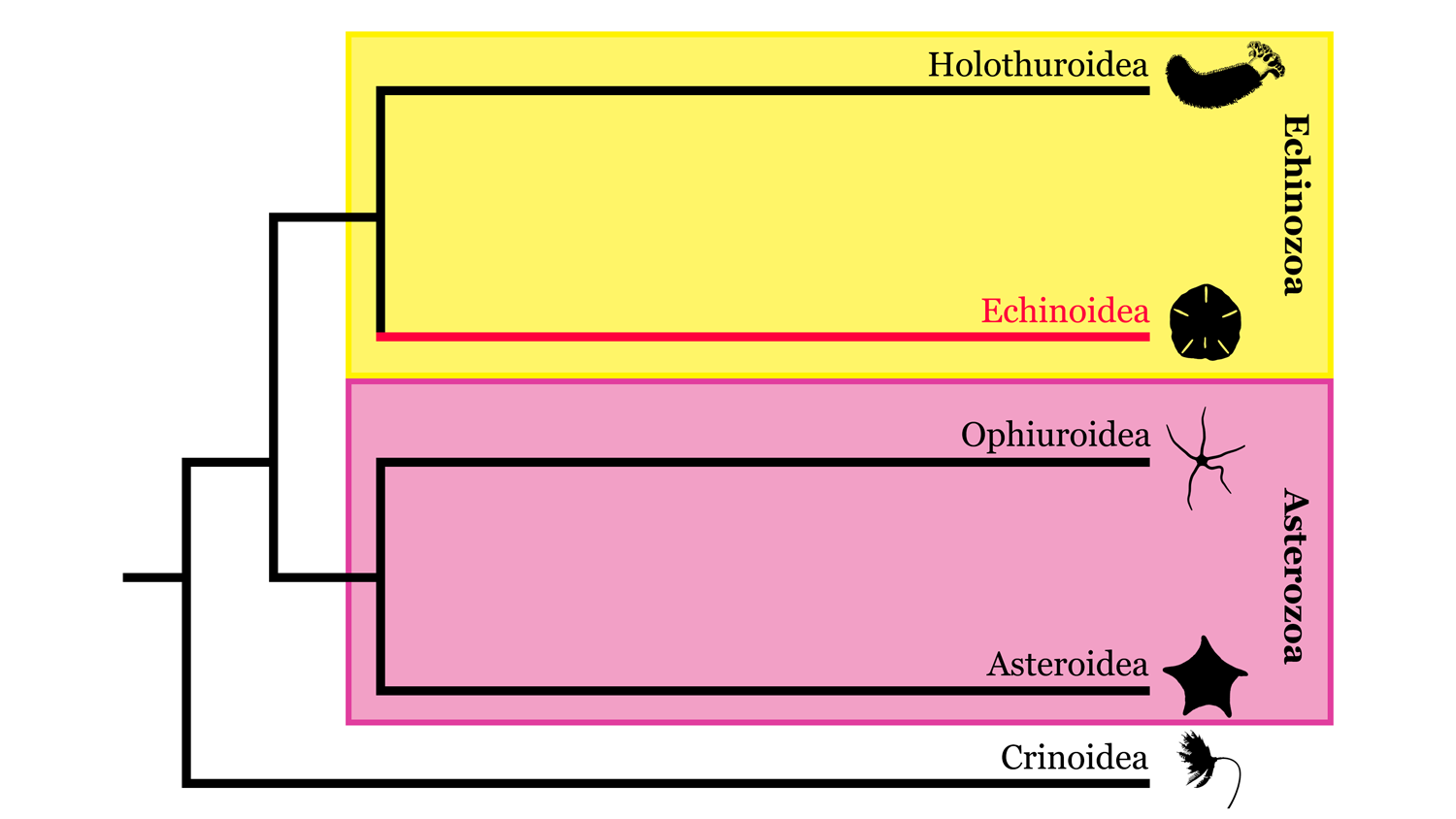
Highly simplified overview of Echinodermata phylogeny based in part on the hypothesis of relationships presented by Reich et al. (2015). Image by: Jaleigh Q. Pier, licensed under a Creative Commons Attribution-Share Alike 4.0 International License.
Colloquially, echinoids take two general forms: regular and irregular. Regular echinoids, which include the sea urchins, tend to be herbivorous (e.g., eating algae, kelp) or carnivorous (e.g., eating bryozoans) and epifaunal (i.e., living on the surface of ocean bottom sediments). The irregular echinoids, which include sand dollars and sea biscuits, tend to be deposit feeders (i.e., eating tiny food particles contained within the sediment) and infaunal (i.e., living buried in ocean bottom sediments). As discussed below, these two groups are readily distinguished by their morphologies and ecologies. As a function of this variety, echinoids encompass one of the most diverse echinoderm classes. Learn more fun facts about echinoids in the video below!
"10 Things You Didn't Know About Sea Urchins" by geobeats (YouTube).
Echinoid morphology
All modern echinoids have a hard calcareous endoskeleton made up of interlocking plates, called a test. In most Paleozoic echinoids, the test was less rigid, and test plates did not interlock as strongly as in modern taxa. Since the end of the Paleozoic, most echinoid tests have been composed by two sets of plates—ambulacral (through which tube feet protrude) and interambulacral. Typically, two rows of each type adjoin each other and alternate between ambulacral and interambulacral areas. Explore the tests of modern and fossil regular echinoids in the 3D models below, which include annotations of key morphological features.
Modern specimen of the purple sea urchin Strongylocentrotus purpuratus from Cornwall, England (PRI 78783). Specimen is from the research collections of the Paleontological Research Institution, Ithaca, New York. Maximum diameter of specimen is approximately 12 cm.
Fossil specimen of Phymosoma texanum from the Cretaceous Fredericksburg Group of Texas (PRI 76726). Specimen from the collections of the Paleontological Research Institution, Ithaca, New York. Specimen is approximately 4.5 cm in diameter.
In living individuals, spines are attached to the test via small ball-in-joint sockets. Because spines are held in place by muscles that rapidly decay after death, they rapidly disarticulate when individuals die. As a consequence, disarticulated echinoid spines are numerous in some portions of the fossil record. In addition to being used for protection from predators, some echinoids use their spines to move along the ocean bottom (watch an echinoid “walk” in the video below). When not using their spines for locomotion, echinoids use their abundant tube feet to move (see anatomical image below).
"How Do Sea Urchins Walk" by Shazaad Kasmani (YouTube).
In regular echinoids (such as shown in the image below), the mouth (or, peristome) is on the bottom, substrate-facing side of individuals and the anus (or, periproct) is located on top. Inside the mouth is a five-part jaw apparatus, called Aristotle’s lantern, which is used to break apart food and to scrape surfaces for algae during grazing.
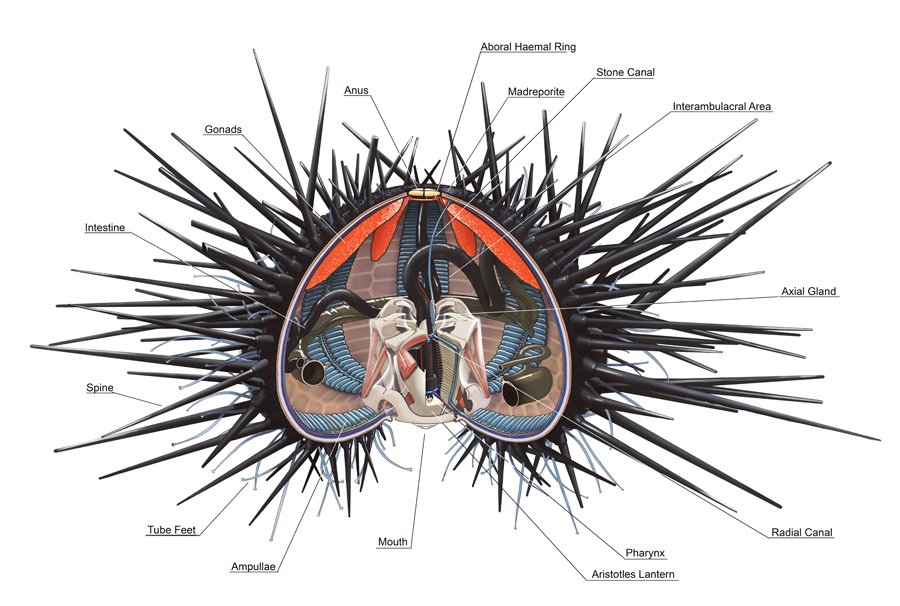
Generalized internal and external anatomy of a regular echinoid, based on the sea urching Arbacia. Image by Alex Ries; Creative Commons Attribution-Share Alike 4.0 International license.
{kind=link}
Given that they commonly feed by excavating sediment and ingesting food particles, irregular echinoids do not possess a lantern and, in many species within this group, the mouth is on the bottom, but the anus is shifted to either the posterior side or bottom of the test. Thus, irregular echinoids are secondarily bilaterially symmetrical in form. Explore the tests of the two fossil irregular echinoids in the 3D models below, which include annotations of key morphological features.
Fossil sand dollar Encope tamiamiensis from the upper Pliocene Tamiami Fm. of Charlotte County, Florida (PRI 44095). From the collections of the Paleontological Research Institution. Maximum length of specimen approximately 7 cm.
Fossil specimen of the irregular echinoid Eupatagus antillarum from the Eocene Ocala Limestone of Levy County, Florida (PRI 76727). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Length of specimen is approximately 7 cm.
In both regular and irregular echinoids, the mouth and anus are connected by a complete digestive system. Both also typically possess a madreporite and gonopores, which are located near the anus. The madreporite controls the pressure in the water vascular system. The gonopores, which are connected to internal gonads, allow for the release of reproductive gametes. Among the Echinoidea, variation exists in many of these features, but most species do follow these general anatomical patterns.
Echinoid diversity and ecology
Echinoids originated during the Ordovician (learn more in the section below on the echinoid fossil record) and are represented in the fossil record by more than 10,300 extinct species. Echinoids have generally increased in diversity moving towards the present, with more than 4,200 species known today. Whereas some groups of echinoids are quite old, like the cidaroids (e.g., pencil urchins; see the 3D model of Eucidaris below) that first evolved in the Permian (~295 million years ago), other groups evolved relatively recently, such as the Clypeasteroids (i.e., sand dollars; see the 3D models of Parascutella below), which originated during the Paleocene (~56 million years ago). The group of echinoids that includes sand dollars, sea biscuits, and heart urchins are generally referred to as irregulars, and comprise the monophyletic infraclass Irregularia. The remaining echinoids, commonly referred to as sea urchins, are known as the regular echinoids. The regular echinoids are a paraphyletic group, but the term is commonly used out of convenience.
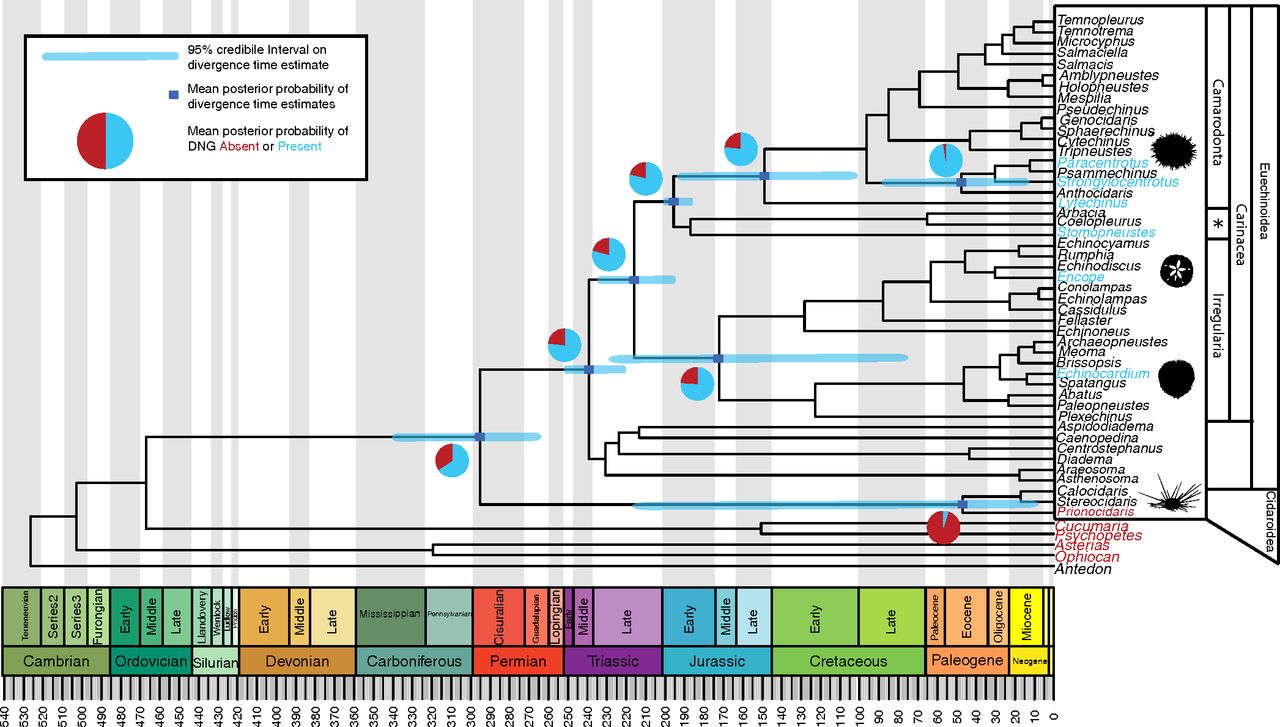
An echinoid phylogeny with estimated times of divergence for major groups. "Fossil-calibrated time tree showing age of clades of crown group echinoids based on one of the 15 alternative topologies resulting from the Bayesian phylogenetic analysis and showing the probability of the presence or absence of the DNG on nodes of interest. Blue bars are 95% CIs on divergence times for nodes. Pie charts show mean PPs that the DNG was present (blue) or absent (red) at particular ancestral nodes in ancestral state reconstructions on 1,300 alternative topologies under priors on transition rates of U(0, 0.01). The taxa in blue have been demonstrated experimentally to use the DNG for micromere specification, whereas the taxa in red do not use the DNG. Taxonomic names represent higher-level echinoid taxonomic groupings. PPs for ancestral state reconstructions are presented in SI Appendix, Table S8. *Indicates Stomopneustoida + Arbacioida. The geological timescale was created in strap (62). Illustrations of spatangoid silhouette, by Hans Hillewaert (vectorized by T. Michael Keesey); cidaroid silhouette, by Didier Descouens (vectorized by T. Michael Keesey); and regular euechinoid, from Frank Förster (based on a picture by Jerry Kirkhart, modified by T. Michael Keesey), are available for use under a Creative Commons Attribution-ShareAlike 3.0 Unported license. Clypeasteroid image, from Michael Site, is available for use under a Creative Commons Attribution-NonCommercial 3.0 Unported license. All images are from Phylopic." Figure and caption from Thompson et al. (2017) in Proceedings of the National Academy of Sciences under the PNAS License to Publish (Copyright (2017) National Academy of Sciences).
Regular echinoids
The regular echinoids (i.e., sea urchins) are a paraphyletic group—due to the position of Irregularia nested within the “regular” echinoids (see phylogeny above)—most easily recognized by their globular shape, with the mouth positioned centrally on the underside and the anus centrally on the top side. With this vertically oriented body plan, urchins do not have a “front” or “back” and move in any direction, rather than front-first. Though they are capable of moving on soft sediments, sea urchins more commonly live(d) on hard substrates, such as reefs and rocks. Many members of this group are herbivorous and, left unchecked by natural predators (e.g., sea stars, otters), can decimate marine vegetation (e.g., kelp forests).
"Can we save kelp forests by eating sea urchins?" by Associated Press (YouTube).
Other urchins are carnivorous, consuming other marine invertebrates like crinoids and bryozoans. In fact, predation by urchins during the Mesozoic likely contributed to the evolution of defensive strategies in these groups, with stalked crinoids evolving to be more mobile and bryozoans evolving enhanced defensive structures (learn more about Bryozoans and Crinoids). Much as they do when consuming vegetation, urchins use their powerful Aristotle’s lantern to latch onto and chew their food. Watch the video below to see an echinoid devour a crinoid.
Watch as a sea urchin pursues and consumes a crinoid. "Sea Urchin Predators Drive Evolution" by Laboratory Equipment (YouTube).
Urchins are often the target of predation themselves, being consumed by sea stars, otters, fish, and even people. Though much of the urchin is hard skeleton and water vascular system, the gonads, of which there are five, are a nutritious, calorie-rich food source. Many species use their long spines as deterrents to predators, with some species even evolving toxins to further avoid being consumed. Urchin spines are so effective at deterring predators that some crab species have co-opted the urchins for a similar function.
"Watch: Carrier Crab Uses Spiny Urchin As Shield | National Geographic" by National Geographic (YouTube)
On a smaller scale, many urchins also possess claw-like structures called pedicellariae (see image below). Urchins use their pedicellariae to remove debris and pests from their surfaces and some species use them to deliver toxins to potential predators. Still other species, like the collector urchin (Tripneustes gratilla), use these grasping structures to cover themselves with small shells and rocks for camouflage.
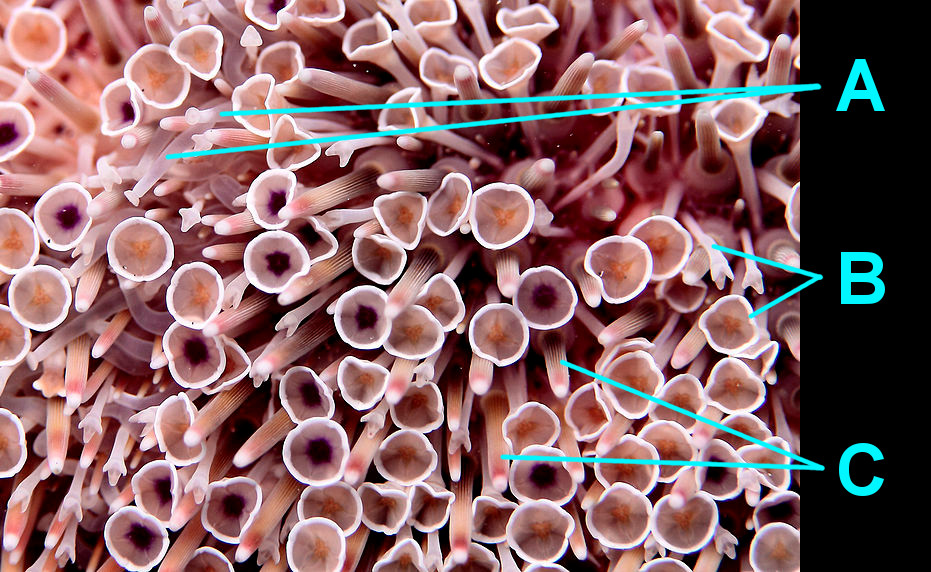
A close up of the modern "flower urchin" showing its pedicellariae (A), tube feet (B), and spines (C). Image by Philippe Bourjon; Creative Commons Attribution-Share Alike 3.0 Unported license.
{kind=link}
Observe the 3D models of other regular urchins below. These urchins have broad, dense spines and are commonly referred to as pencil urchins. These urchins are considered more primitive because their morphology more closely resembles ancestral echinoids and, as shown in the phylogeny above, pencil urchins, like the cidaroids, split from other echinoids during the Permian.
Modern specimen of the slate pencil sea urchin Heterocentrotus mamillatus. Specimen is from the research collection of the Paleontological Research Institution, Ithaca, New York. Maximum diameter of specimen is approximately 15 cm. Model by Emily Hauf.
Modern specimen of the echinoid Eucidaris tribuloides from near Key West, Florida (PRI 13410). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Specimen is approximately 7.5 cm in diameter. Model by Emily Hauf.
Irregular echinoids
The irregular echinoids are a monophyletic group, called Irregularia, that includes common forms like sand dollars (order Clypeasteroida; see the Encope 3D model above), sea biscuits (order Clypeasteroida; see the Clypeaster 3D model below), and heart urchins (order Spatangoida; see the 3D model of Echinocardium below). Irregular echinoids are morphologically distinct from the regular echinoids. In the irregulars, the mouth and anus are shifted toward opposite sides of the test (to varying degrees), rather than being centrally located on the top (anus) and bottom (mouth) of the test. Additionally, many irregulars have a flattened, rather than globose appearance, and have much shorter spines compared to regular echinoids.
Modern specimen of the sea biscuit Clypeaster reticulatus from the Indo-Pacific Ocean (T-1818). Specimen is from the teaching collection of the Paleontogical Research Institution, Ithaca, New York. Length of specimen is approximately 13 cm. Model by Emily Hauf.
Fossil specimen of the heart urchin Echinocardium orthonotum from the Miocene of Maryland (PRI 503). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Length of specimen is approximately 5 cm.
The morphological changes that led from the bulbous regular form to the flattened form of irregular echinoids reflect their ecology. Unlike regular echinoids, many irregulars are detritivorous and infaunal. When they feed, irregulars move through the sediments and consume tiny particles of organic material. Their flattened form and reduced spines are more conducive to burrowing than the globose, spiny form of regular echinoids. In addition to advantages for their detritivorous feeding, the infaunal life mode of irregulars serves to protect them from predators. In the absence of large spines and pedicellariae for defense, burrowing provides a refuge for irregulars by concealing them from their predators.
Learn more about sand dollars and how they feed. "A Sand Dollar's Breakfast is Totally Metal" by Deep Look (YouTube).
Watch as a heart urchin moves along the seafloor and buries itself using its spines. "Heart Unchin" by Sea School (YouTube).
Today, irregular echinoids are highly diverse and live in most marine environments, despite the relatively recent evolution of many groups in Irregularia. Watch the video below to review some of the details about the irregular echinoids.
Learn more about the life of a sand dollar. "Facts: The Sand Dollar" by Deep Marine Scenes (YouTube).
Echinoid fossil record
Echinoids first appear in the fossil record in the Upper Ordovician, approximately 460 million years ago. Their time of first appearance coincides with the first appearance of holothurians, the class of echinoderms to which echinoids are most closely related. The other three extant echinoderm classes evolved earlier. Echinoids are relatively rare in the Paleozoic fossil record. This rarity is likely related to the fragility of the tests of early echinoids. Compared to the tests of many modern echinoids, Paleozoic echinoid tests were less rigid, and plates were held together by ligaments. After dying, these tests rapidly disarticulated, leaving a poor fossil record.
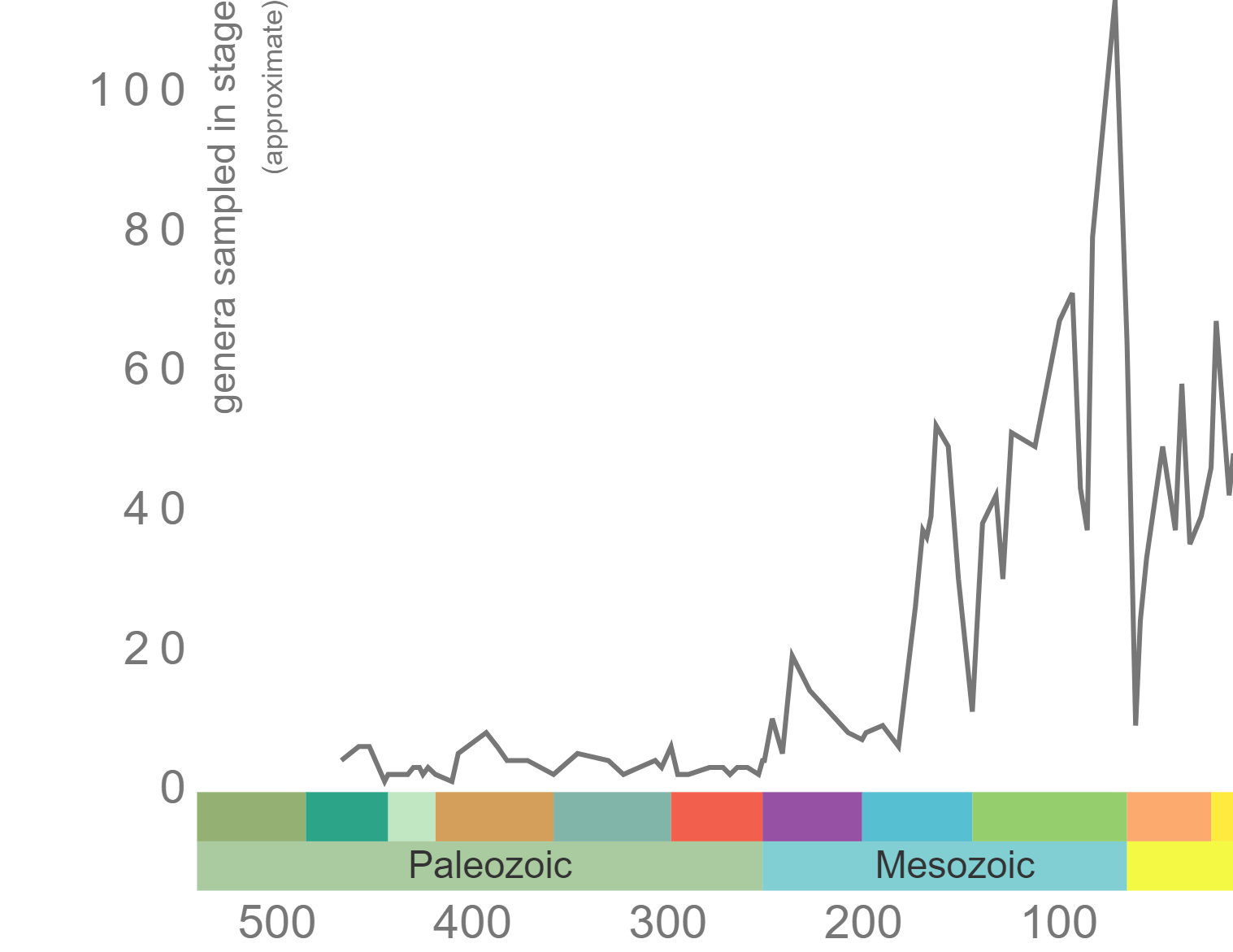
Phanerozoic genus-level diversity of Echinoidea (graph generated using the Paleobiology Database Navigator).
During the Devonian, regular echinoids began to resemble their modern form. It is during this time that the first pedicellariae and coverings of large spines are evolved, suggesting an increase in predatory pressure. The end-Permian mass extinction nearly eliminated the echinoids, as potentially only one group, the cidaroids, survived. Thus, cidaroids may be ancestral to all living echinoids. Moving into the Mesozoic, the fossil record of echinoids improves, potentially due to the evolution of a more rigid, preservable test.
Fossil specimens of the echinoid Archaeocidaris immanis from the Pennsylvanian Winchell Limestone of Brown County, Texas (PRI 45520). Specimen is on display at the Museum of the Earth, Ithaca, New York. Longest dimension of rock is approximately 21 cm. Model by Emily Hauf.
Fossil cidaroid sea urchin Balanocidaris marginata from the Jurassic of France (PRI 78787). Specimen is from the research collections of the Paleontological Research Institution, Ithaca, New York. Maximum diameter of specimen is approximately 7 cm. Model by Emily Hauf.
It was not until the Late Triassic that echinoids recovered from the end-Permian extinction and began to diversify. Among this diversity were the cidaroids (i.e., pencil urchins), which first evolved during the Permian; the pedinoids, which were diverse and abundant during the Jurassic, with only a single deep-water genus, Aulosonta, surviving today; and eventually, in the Jurassic, the carinaceans, which include many of today’s regular echinoids. This increase in diversity also coincides with the first feeding traces produced by the Aristotle’s lanterns mouth component, and development of morphologically diverse pedicellariae in the echinoids. Both innovations likely reflect the overall increases in predation—on and by echinoids—and competition during this time, often referred to as the Mesozoic Marine Revolution.
Interestingly, after the Mesozoic, the fossil record of regular echinoids is relatively poor. As discussed by Smith and Kroh (2013), the paucity of regular echinoids is likely a consequence of their transition to hard substrates, where preservational potential is low. Starting in the Middle Jurassic and continuing through the Cenozoic, irregular echinoids are abundant in the fossil record from soft sediment habitats, essentially replacing the regular echinoids in those habitats. Because many irregular echinoids are infaunal, living within the sediments, their excellent fossil record is no surprise—when they die, they are already buried and on their way to becoming fossils. These early irregular echinoids were largely of the heart urchin variety, as it was not until the Paleocene that sand dollars (i.e., clypeasterines, scutellines) evolved. From the Eocene onward, sand dollar forms became common, and they remain so today. Check out some fossil irregular echinoids in the 3D images below.
Fossil specimen of the heart urchin Schizaster sp. from the Eocene La Meseta Formation of Seymour Island, Antarctica (PRI 58941). Specimen is from the research collections of the Paleontological Research Institution, Ithaca, New York. Length of specimen is approximately 5.5 cm. Model by Emily Hauf.
Fossil sand dollars belonging to the species Parascutella paulensis from the Miocene of Bordeaux, France (PRI 78782). Specimen is from the research collection of the Paleontological Research Institution, Ithaca, New York. Length of rock is approximately 20 cm. Model by Emily Hauf.
Conglomerate rock composted mostly of the tests of sand dollars (species unidentified); specimen is from the Neogene of Santa Cruz County, California (PRI 81257). Specimen is from the research collections of the Paleontological Research Institution, Ithaca, New York. Length of rock is approximately 13.5 cm. in length. Model by Emily Hauf.
References and further reading:
Boardman, R.S., Cheetham, A.H., and Rowell, A.J. 1987. Fossil Invertebrates. Blackwell Scientific Publications. 713 pp.
Kroh, A. 2020. Phylogeny and classification of echinoids. In Developments in Aquaculture and Fisheries Science (Vol. 43, pp. 1-17). Elsevier.
Kroh, A., and A. B. Smith. 2010. The phylogeny and classification of post-Palaeozoic echinoids. Journal of Systematic Palaeontology, 8: 147-212.
Littlewood, D. T. J., and A. B. Smith. 1995. A combined morphological and molecular phylogeny for sea urchins (Echinoidea: Echinodermata). Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 347: 213-234.
Nichols, D., 1967. Echinoderms. Hutchinson University Library, London.
Smith, A. B. 2009. Sea urchins (Echinoidea). In: The Timetree of Life, eds. S. B. Hedges and S. Kumar. Oxford University Press, Oxford. pp. 302-306.
Thompson, J. R., E. M. Erkenbrack, V. F. Hinman, B. S. McCauley, E. Petsios, and D. J. Bottjer. 2017. Paleogenomics of echinoids reveals an ancient origin for the double-negative specification of micromeres in sea urchins. Proceedings of the National Academy of Sciences, 114: 5870-5877.
Usage
Unless otherwise indicated, the written and visual content on this page is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. This page was written by Jansen A. Smith and Jaleigh Q. Pier. See captions of individual images for attributions. See original source material for licenses associated with video and/or 3D model content.
