


Chapter contents:
Echinodermata
–– 1. Exclusively Fossil Taxa ←
–– 2. Crinoidea
–– 3. Asteroidea
–– 4. Ophiuroidea
–– 5. Echinoidea
–– 6. Holothuroidea
This page was written by Jansen Smith. It was last updated on June 22, 2020.
Above image: Specimens of the eocrinoid, Gogia, from the Middle Cambrian of Utah (USA). Image by James St. John; Creative Commons Attribution 2.0 Generic license.
Fossil Taxa Snapshot
- Examples: Blastoidea, Diploporita, Eocrinoidea, Edrioasteroidea, Helicocystoidea, Stylophora
- Ecology: Marine; filter feeders, herbivores, deposit feeders
- Key features of group: multi-plated calcium carbonate skeletons with stereom, water vascular system
- Diversity: ~28 classes
- Fossil record: Cambrian to Permian
Overview
The earliest echinoderms evolved a variety of forms that make them almost unrecognizable as members of this phylum. For example, many of the species in the image below exhibit bilateral symmetry rather than the pentaradial symmetry that characterizes this phylum. Others (e.g., Helicocystoidea) appear to be transitional forms between the less familar forms and the echinoderms we see commonly today. In the sections that follow, several classes belonging to each of the three morphological groups are introduced—non-radial; stalked radial; and pentaradial attached and mobile—providing an entry-level survey of the diversity of early echinoderms (the taxonomy, and inclusion of classes, is based on dataset from the recent work by Deline et al. 2020 deposited with Dryad).
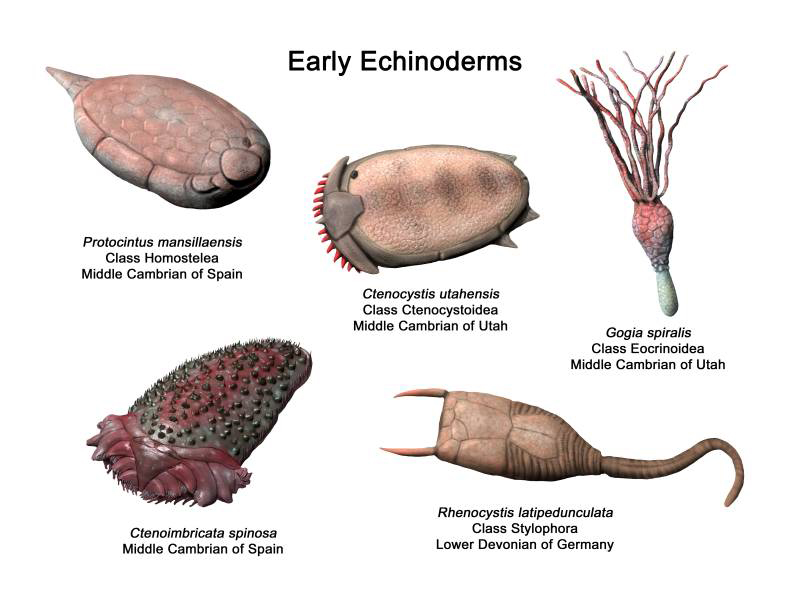
Non-radial forms
The non-radial forms of echinoderms are almost non-recognizable as echinoderms because they do not exhibit pentaradial symmetry, which is a defining feature of the phylum. Five classes are included in this group: Ctenocystoidea, Ctenoimbricata, Cincta, Stylophora, and Soluta. Even amongst these classes, there is relatively little overlap in morphology, as each class is relatively distinct. The classes Ctenocystoidea, Ctenoimbricata, and Stylophora are highlighted below.
Ctenocystoidea
Ctenocystoids, known from the Cambrian and Ordovician, are distinguishable from many other echinoderms by their bilateral symmetry, similar to the ancestors of the echinoderm phylum, which remain unknown but are thought to be bilaterians. As discussed by Rahman et al. (2015), this potential resemblance to the hypothetical, bilateral ancestral form has led some researchers to argue that this class is one of the most ancient echinoderm lineages. Others have suggested that, instead of retaining the ancestral body plan, ctenocystoids secondarily evolved their bilateral symmetry—implying that the direct ancestor of ctenocystoids likely had a body plan with pentaradial symmetry. The exact phylogenetic position of this class remains a point of debate.
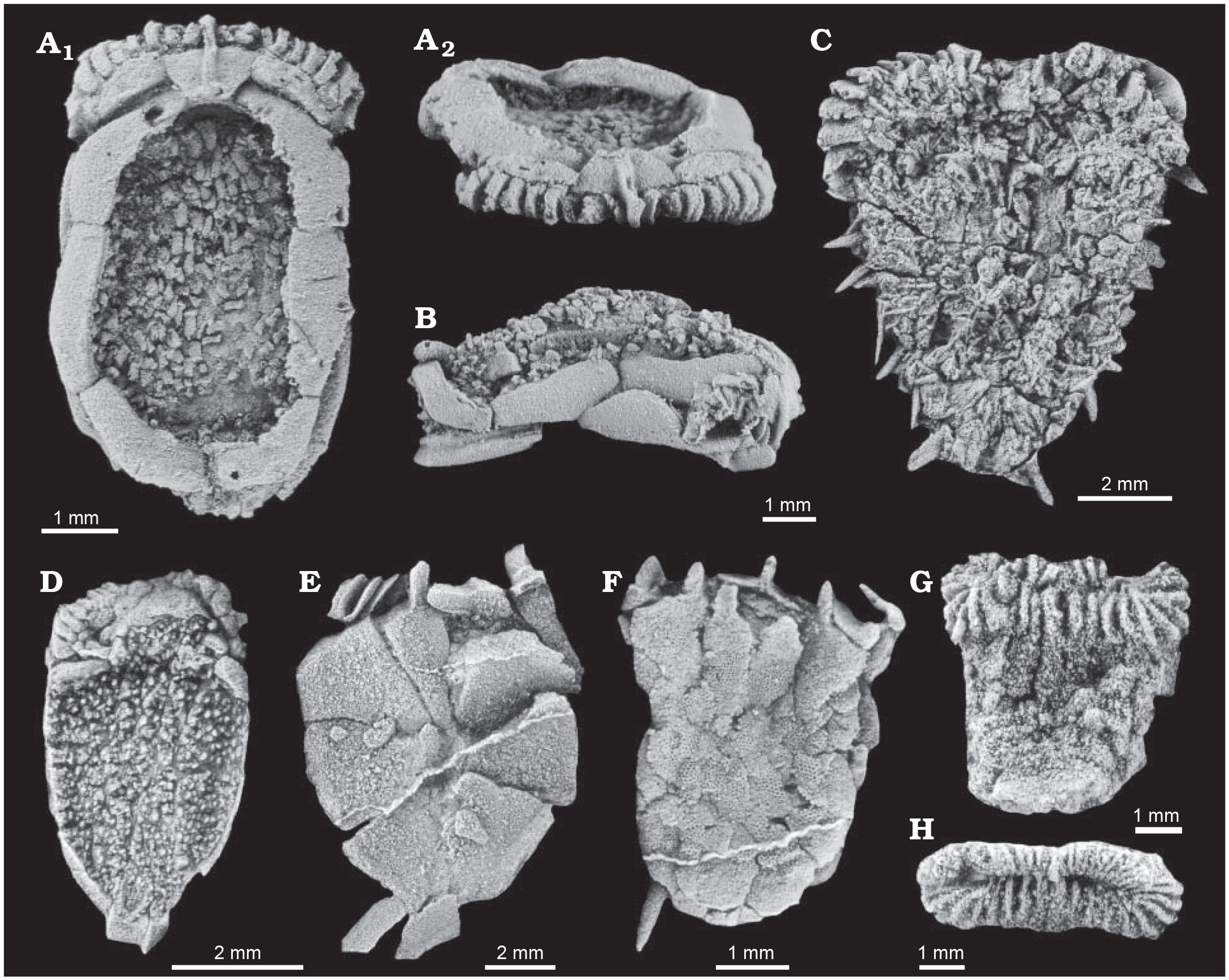
A collection of ctenocystoids from the Cambrian. Note the scale bars indicating the small size of these animals. "Representative Cambrian ctenocystoid echinoderms. A, B. Ctenocystis utahensis Robison and Sprinkle, 1969 from Cambrian Series 3 of the United States. USNM 163252 in dorsal (A1) and anterior (A2) views; USNM 595079 in right lateral view (B). C. Undescribed ctenocystoid (Ctenocystoid gen. et sp. nov 1 in Smith et al. 2013) from Cambrian Series 3 of Morocco. NHMUK EE 15428 in dorsal view. D. Courtessolea moncereti Domínguez Alonso, 2004 from Cambrian Series 3 of France. MNHN F.A45783 in dorsal view. E. Courtessolea sp. from Cambrian Series 3 of Spain. MPZ 2009/1234b in ventral view. F. Undescribed ctenocystoid (Ctenocystoid gen. et sp. nov 2 in Smith et al. 2013) from Cambrian Series 3 of Morocco. NHMUK EE 15317 in ventral view. G, H. Jugoszovia archaeocyathoides Dzik and Orłowski, 1995 from Cambrian Series 3 of Poland. ZPAL Ec1/9 in ventral view (G); ZPAL Ec1/1 in anterior view (H). All specimens are latex casts whitened with ammonium chloride sublimate." Figure and caption from Rahman et al. (2014) in Acta Palaeontologica Polonica; Creative Commons Attribution 4.0 International license.
Ctenocystoids were likely mobile, with forms like Conollia reconstructed to be burrowers, or at least semi-infaunal (see below). If it was semi-infaunal, Conollia likely consumed small organic particles from the surface of ocean bottom sediments, which is a commonly suggested life habit for ctenocystoids. If infaunal, Conollia and other ctenocystoids were instead likely deposit feeders, extracting food particles from buried sediments.
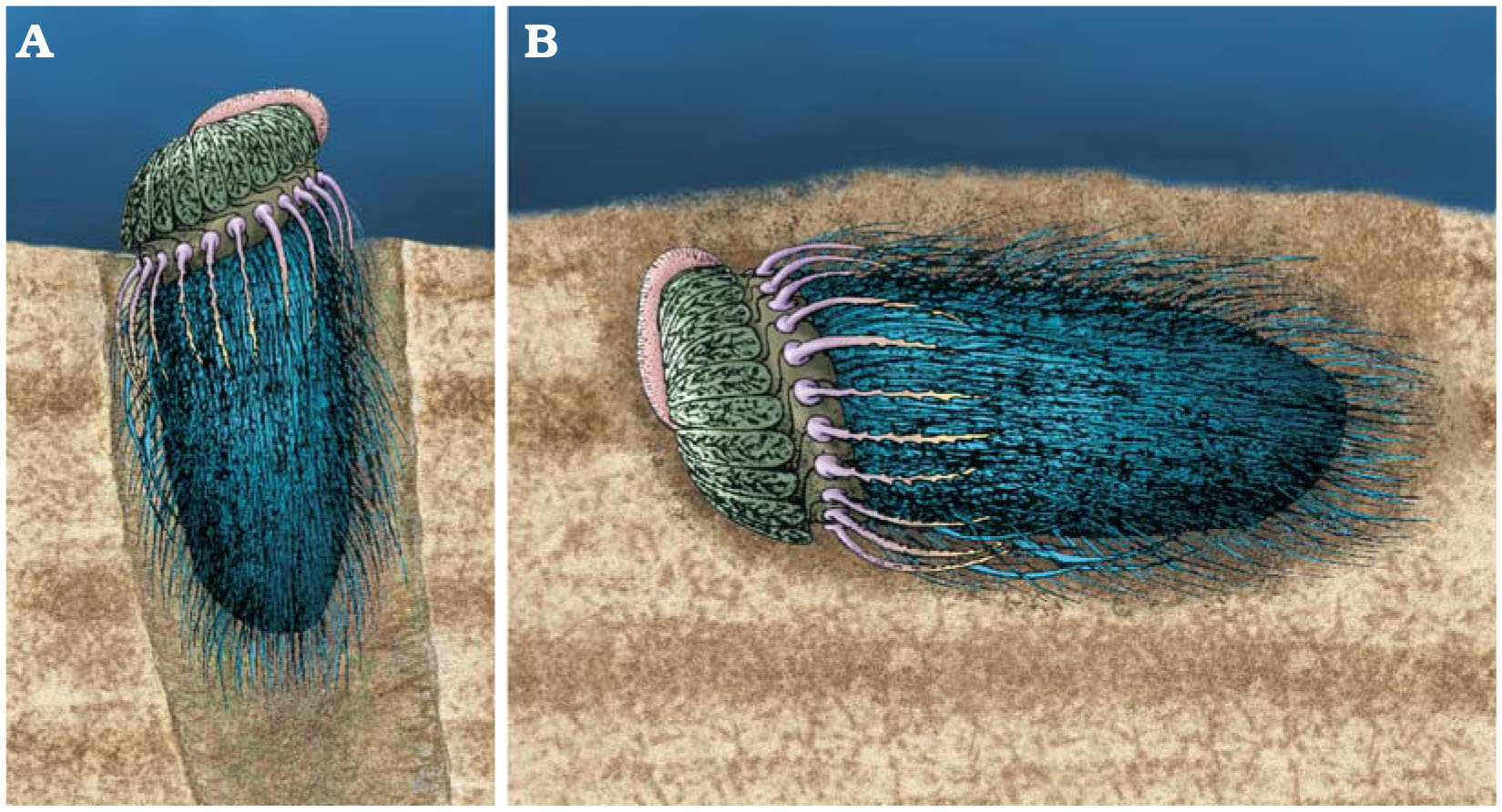
"Reconstructions of two possible modes of life of Conollia. A. A semi-infaunal mode of life. B. An infaunal mode of life. Adapted from Domínguez Alonso (2004). "Figure and caption from Rahman et al. (2014) in Acta Palaeontologica Polonica; Creative Commons Attribution 4.0 International license."
Ctenoimbricata
The Cambrian Ctenoimbricata, like the ctenocystoids, exhibit bilateral symmetry. Unlike the ctenocystoids, however, this body plan is thought to be ancestrally derived in Ctenoimbricata, suggesting that this group may be basal to all other known echinoderms. Moreover, the discovery and description of this group by Zamora et al. (2012) supported the idea that ancestral echinoderms had bilateral symmetry, and the characteristic pentaradial symmetry within the phylum evolved some time later. As shown in the reconstruction below, ancestral echinoderms like Ctenoimbricata were likely deposit feeders rather than suspension feeders, though this latter life mode would become very common among echinoderms and still is today.
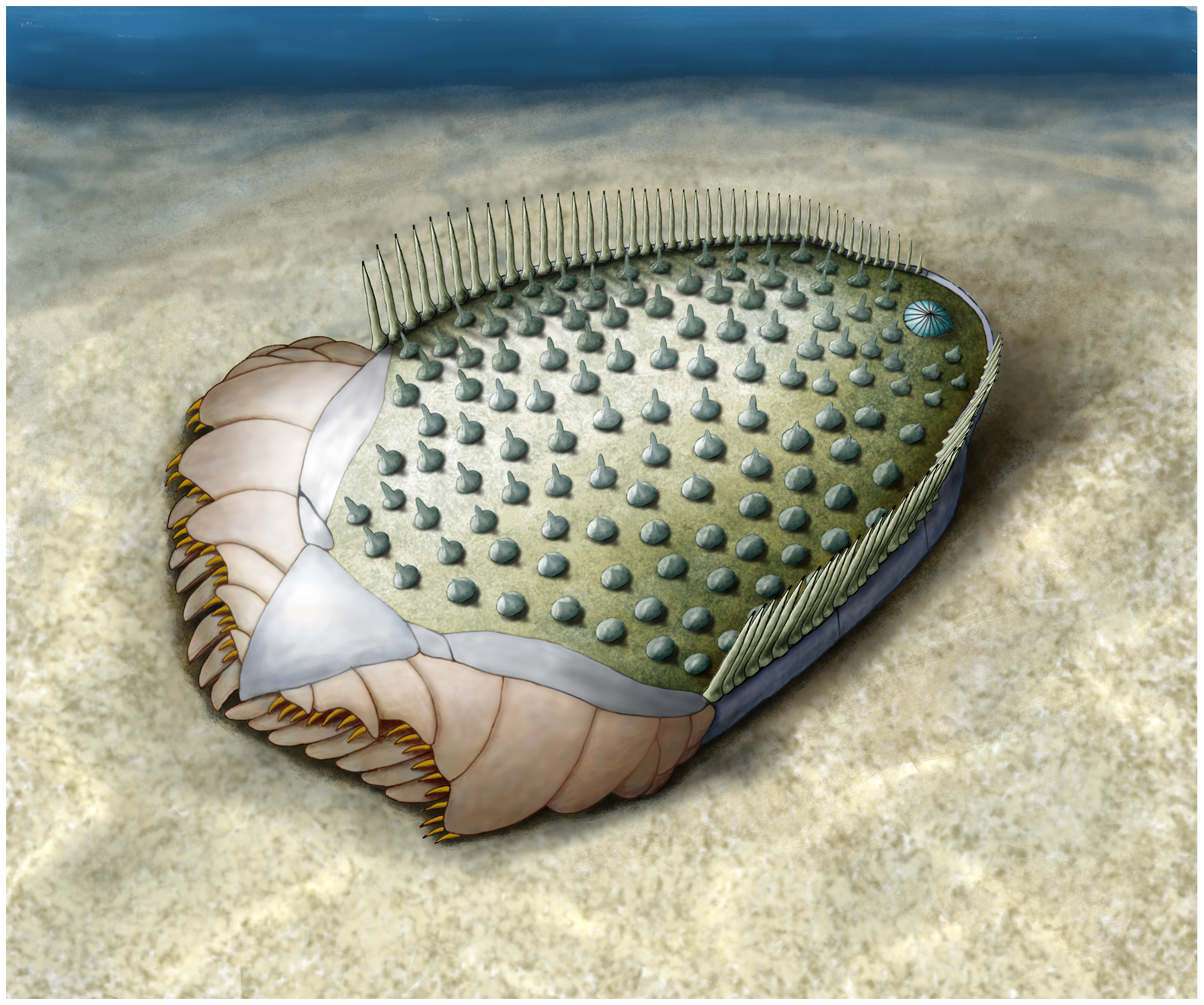
"Reconstruction of Ctenoimbricata spinosa gen. et sp. nov." Figure and caption from from Zamora et al. (2012) in PloS ONE; Creative Commons Attribution-Share Alike 4.0 International license.
Stylophora
The strange appearance of the Stylophora—occurring from the Cambrian to Carboniferous—has led to controversy in reconstructions of how the animals in this class lived, and where they should be placed on the tree of life. Though there is agreement today that this class belongs in the Echinodermata, earlier assignments placed the lineage as primitive chordates—the phylum that includes us. Placement with the chordates was based in large part on the hypothesis that the long appendage of the stylophorans contained a notochord. Others suggested that this appendage was analogous to the stalk of other echinoderm groups (e.g., crinoids, blastoids) and used to anchor the stylophorans in the sediment. These hypotheses have been rejected, as scanning electron microscopy by Lefebvre et al. (2019) showed the presence of a longitudinal canal with projections coming from it, much like the water vascular system in other echinoderms. These authors concluded that the appendage probably functioned like the arm of a sea star and may have been used to capture food. With the combined presence of a water vascular system and ossicles with stereom microstructure—a specific type of calcium carbonate used by echinoderms—there can be little doubt that the Stylophora belong in Echinodermata.
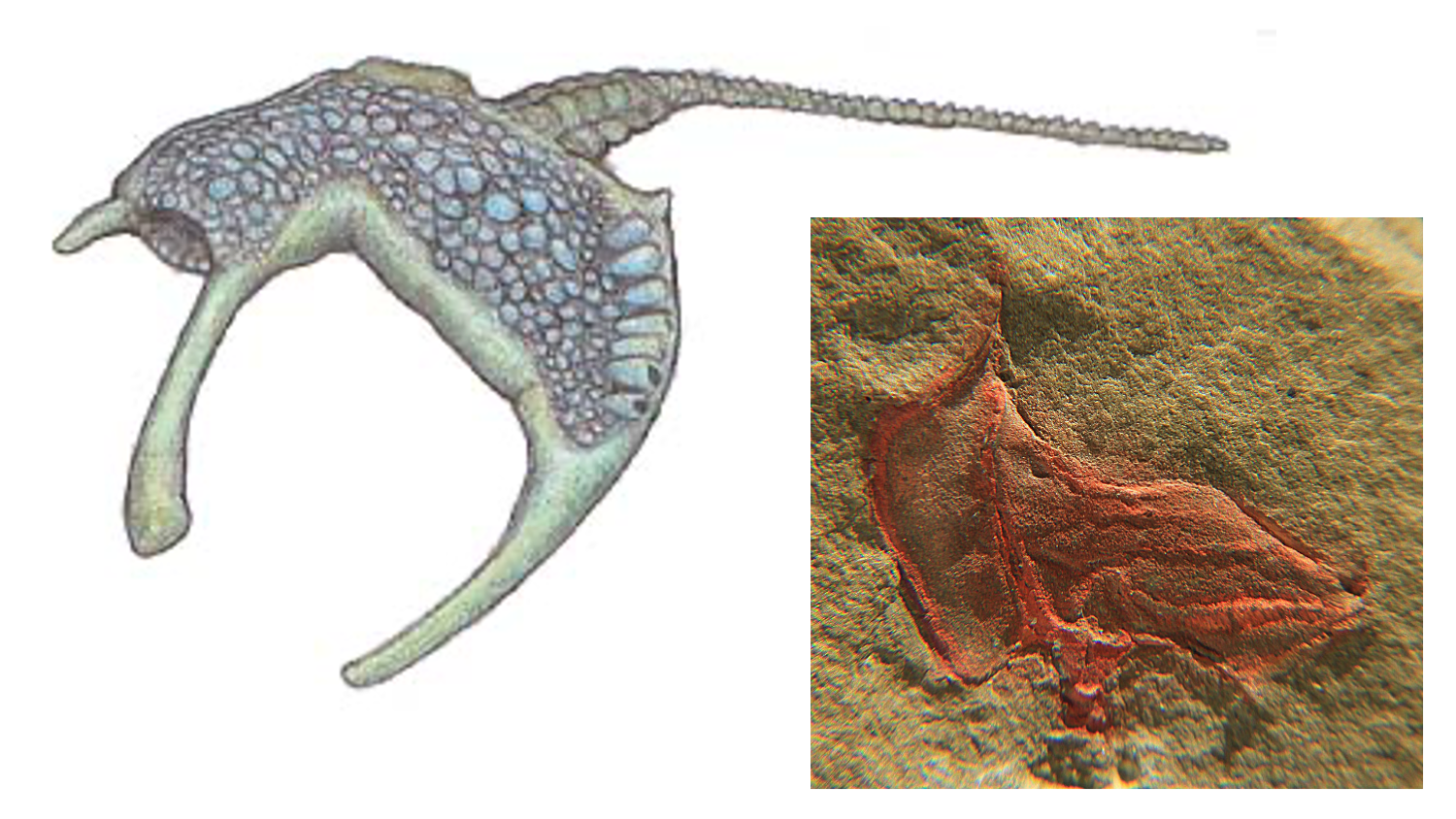
A reconstruction of the Ordovician stylophoran, Cothurnocystis (left), and a fossil of the same species from Morocco. Left image by Dwergenpaartje; Creative Commons Attribution-Share Alike 3.0 Unported license. Right image by Goombasaurusrex; Creative Commons Attribution-Share Alike 3.0 Unported license.
{kind=link}
Stalked radial forms
The stalked radial echinoderms include two forms, as defined by Deline et al. (2020), arguably forming a clade called Pelmatozoa. One group includes the crinoids, and the other (i.e., blastozoans) contains a variety of stalked classes, most notably the blastoids. One of the major differences between these groups is in the structure used to capture food. The group including blastoids used brachioles, whereas the crinoids use arms covered with tube feet. As a living class, crinoids are covered on their own page. The stalked radial classes, which are often distinguished from each other based on structural differences in their water vascular systems, include Blastoidea, Diploporita, Eocrinoidea, Helicocystoidea, and Rhombifera, among others (e.g., Parablastoidea, Paracrinoidea).
Blastoidea
Blastoids, also ocassionally called “sea buds,” get their name from the Greek word "blastos," meaning “bud-like”. Similar to crinoids, they were stalked suspension feeders, but used brachioles (appendages thought to lack the water vascular system) to capture food particles. Blastoids were a common component of crinoid dominated shallow-water communities throughout the Paleozoic. In some assemblages from deep-water environments, evidence suggests they may occassionally have been the numerically dominant echinoderms.

Photograph of a model reconstruction of what blastoids could have looked like in a Carboniferous setting. On display at the Carnegie Museum of Natural History in Pittsburgh, Pennsylvania. Image by: James St. John (Wikimedia Commons; Creative Commons Attribution-ShareAlike 2.0 Generic License)
In blastoids, the calcitic endoskeleton is referred to as a "theca." The theca can be separated into three different types of calcite plates. Deltoid plates (also called “orals”) form the top of the theca near the mouth, radial plates make up the sides, and basal plates form the bottom, which attaches to the stem. Similar to crinoids, blastoids had a stem made up of stacked calcareous disks called columnals, which together form the column. The end of the column anchored to the substrate with a root-like structure. Also, like crinoids, there are some species of blastoid that lacked a column altogether. Click on the 3D models below to explore two species from the blastoid genus Pentremites.
Fossil specimen of the blastoid Pentremites godani from the Mississippian Paint Creek Formation of St. Clair County, Illinois (PRI 70771). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Specimen is approximately 2 cm in length.
Fossil blastoid Pentremites robustus from the Mississippian of Crawford County, Indiana (PRI 44909). Specimen is on display at the Museum of the Earth, Ithaca, New York. Length of specimen is approximately 4.5 cm. Model by Emily Hauf.
The bud-shaped theca has five ambulacra that aided in shuttling nutrients to the mouth, was was located in the center of the topward side. In the middle of each ambulacrum are the ambulacral grooves, canals that facilitated the movement of nutrient rich water from the structures toward the mouth. Each ambulacral groove was supported by a lancet plate, which were sometimes covered by additional small plates. Each food groove would have been lined with thin, string-like brachioles that captured food through suspension feeding. Next to the ambulacral food grooves were lines of hydrospire pores, which are thought to be where water was taken in for respiration, before being expelled through the spiracle, a pore near the mouth. Hydrospire pores are endothecal (inside the theca) structures, where respiration took place within internal canals. Both hydrospire pores and spiracles have been important for distinguishing between different clades and groups of blastoids.

A selection of blastoids drawn by Ernst Haeckel in Kunstformen der Natur (1904), plate 80: Blastoidea (public domain). Note, the three specimens along the edges of the lower right quadrant are no longer classified as blastoids.
{kind=link}
Diploporita
Diploporitans are named for their respiratory structures, diplopores. Each diplopore has two pores, and many diplopores may occur on the same plate. There is variation in diplopores, particularly in how pores connect. In some groups, the two pores are connected by a single canal, whereas in other groups a complex of canals runs between each set of pores—the specimen in the image below is an example of the latter. This class first appeared in the fossil record during the Ordovician and became speciose, with some estimates of diversity in excess of 175 species, before dwindling to extinction by the end of the Devonian. Despite their diversity, the diploporitan fossil record is relatively incomplete and many species are represented by only a few, potentially incomplete specimens. Paucity notwithstanding, recent work by Sheffield and Sumrall (2019) has confirmed that this group is likely polyphyletic, with the diplopore respiratory structure arising multiple times, independently, in Echinodermata. You can learn more about diploporitans in this article by Dr. Sarah Sheffield.
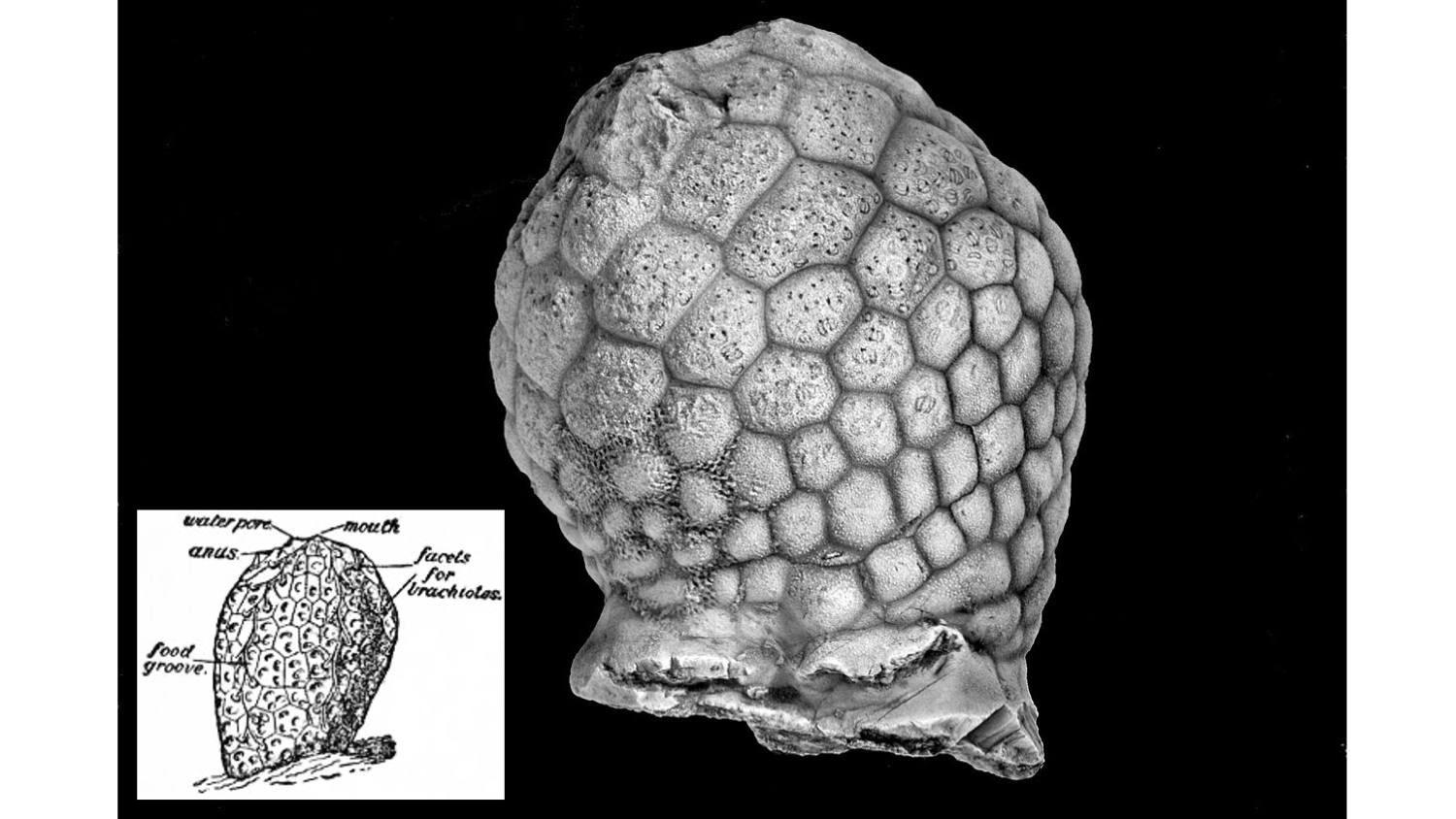
A Silurian diploporitan, Paulicystis sparsus, with an inset showing diploporitan anatomical features on an idealized specimen. Note the canals exposed on the upper half of the specimen in the larger image, which would have connected the diplopores in the living animal. Larger image by Ssheffi2; Creative Commons Attribution-Share Alike 4.0 International license. Inset image from Encyclopedia Britannica (1911; public domain).
{kind=link}
{kind=link}
Eocrinoidea
The eocrinoids were among the first common echinoderms in ocean bottom communities of the Cambrian, though recent work by Nardin et al. (2017) and Nohejlova et al. (2019) suggests the group is not monophyletic. The earliest eocrinoids had very minimal stalks, or simply a holdfast (such as in the right-side image below), and eventually evolved more pronounced stalks (such as in the left-side image below). By elevating the cup and feeding appendages higher into the water column, the eocrinoids were likely able to exploit a broader range of food resources. This group is united by the presence of epispire respiratory structures, which are pores running along the sutures of the skeletal plates. These pores can range in shaper from circular to elliptical to triangular and likely evolved more than once. As it is now thought that “Eocrinoidea” is not a single echinoderm clade, researchers are working to understand the phylogenetic relationships of taxa previously assigned to this group.

A Cambrian fossil eocrinoid, Gogia (left), and a reconstruction of an ocean bottom community, including Gogia (right). Left image by Kevmin; Creative Commons Attribution-Share Alike 3.0 Unported license. Right image by Stanton F. Fink; GNU Free Documentation License.
{kind=link}
{kind=link}
Though their name means “dawn crinoid” and their morphologies are generally similar, this class is not ancestral to the crinoids. As noted above, crinoids use arms and tube feet to feed but blastozoans, like Eocrinoidea, used distinctly different appendages, called brachioles. Given their early appearance in the record and the similarities in structures like brachioles, it has been suggested that the eocrinoids are ancestral to other groups, such as the diploporitans, blastoids, and rhombiferans. It remains challenging to evaluate these potential relationships, however, because many of these groups, Eocrinoidea included, are no longer considered monophyletic.
Fossil eocrinoid Gogia sp. from the Cambrian Spence Shale of Utah. Specimen is on display at the Museum of the Earth, Ithaca, New York. Length of specimen is approximately 8 cm. Model by Emily Hauf.
Helicocystoidea
The helicocystoids are likely among the most primitive of the stalked, radial classes and may represent a transitionary form from primitive bilateral echinoderms. Helicocystoids, such as the Helicocystis moroccoensis featured in the video below, possessed a very short stalk attached to a small cup, reminiscent of the cup in other echinoderm groups (e.g., crinoids). Above the basal cup were skeletal plates, arranged in a spiral, with inset ambulacra. Notably, there are five of these ambulacra, distinguishing this class from other similar, but more primitive forms, such as the Helicoplacoidea. Like in modern echinoderms, the five ambulacra were likely used to transport food captured from the water column into the mouth, which was located on the top of the animal. As suggested by the name, these ambulacra were arranged spirally around the body of the animal, giving it a distinct helical appearance.
Learn more about the helicocystoid, Helicocystis moroccoensis, in this video, "Fossil of 500-Million-Year-Old, Shape-Altering Creature Uncovered in Morocco," by Slate (YouTube).
Rhombifera
The rhombiferans lived from the Ordovician to the Devonian and were epifaunal suspension feeders, like many of the stalked radial blastozoans. Food grooves within the brachioles of rhombiferans were very narrow, suggesting a specialization for feeding on microplankton and other small food particles. It was suggested by Sprinkle et al. (1995) that these animals may have thrived in environments with muddy, nutrient-rich substrates, where resuspension of this bottom layer likely provided ample food.

Examples of an amphorid rhombiferan (left) and Caryocrinites ornatus (right). Left image by Ernst Haeckel (public domain). Right image by James St. John; Creative Commons Attribution 2.0 Generic license.
_1_(17199794079).jpg){kind=link}
As suggested by the name, this class of echinoderms often exhibits rhomb-like features, which were created by the arrangement of infoldings and respiratory pores in the thecal wall. Though the rhombs were once thought to be a synapomorphy of the class, several unique rhomb forms exist, suggesting the pattern arose in multiple groups independently. Based on the differences in how the rhombs arose, it has been suggested that Rhombifera includes multiple clades and may not be monophyletic. In some groups of glyptocystitoid rhombiferans, the number of ambulacra were reduced from the characteristic five, to four, three, two, or one food grooves. When these reductions occurred, they were often associated with increased branching in the remaining ambulacra, suggesting no loss of function due to the reduction. Check out the 3D model below to explore some of these rhombiferans features.
Fossil specimen of the rhombiferan Caryocrinites ornatus from the Silurian of Lockport, New York (PRI 76779). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Length of specimen is approximately 4.5 cm. Model by Emily Hauf.
Pentaradial attached and mobile forms
This group of echinoderm forms includes four of the five living echinoderm classes (i.e., Asteroidea, Echinoidea, Holothuroidea, and Ophiuroidea), and you can learn more about them on their own pages in this chapter. Also included in this group are stationary, attached (e.g., Cyclocystoidea, Camptostromoidea, Edrioasteroidea) classes and a variety of other free-living, mobile (e.g., Ophiocistoidea, Somasteroidea) classes. In the sections that follow, we will take a closer look at two of the attached classes, Cyclocystoidea and Edrioasteroidea, and one of the mobile classes, Ophiocistoidea.
Cyclocystoidea
Cyclocystoids are a relatively rare and poorly known echinoderm class that lived from the Ordovician to the Carboniferous. Typically, cyclocystoids are no more than a centimeter in diameter and are preserved as rings of plates, or individual plates (see the image below). Because they are typically preserved incomplete—missing the components within the ring structure—the soft tissues of this class are not known and remain the subject of debate. The life habitat of this enigmatic group is likewise poorly known. Some have suggested that cyclocystoids were mobile deposit feeders, but others have argued that they lived a stationary life on the sea floor as suspension feedings. Until more specimens, and especially well-preserved specimens, are found, much about this group will remain unknown.
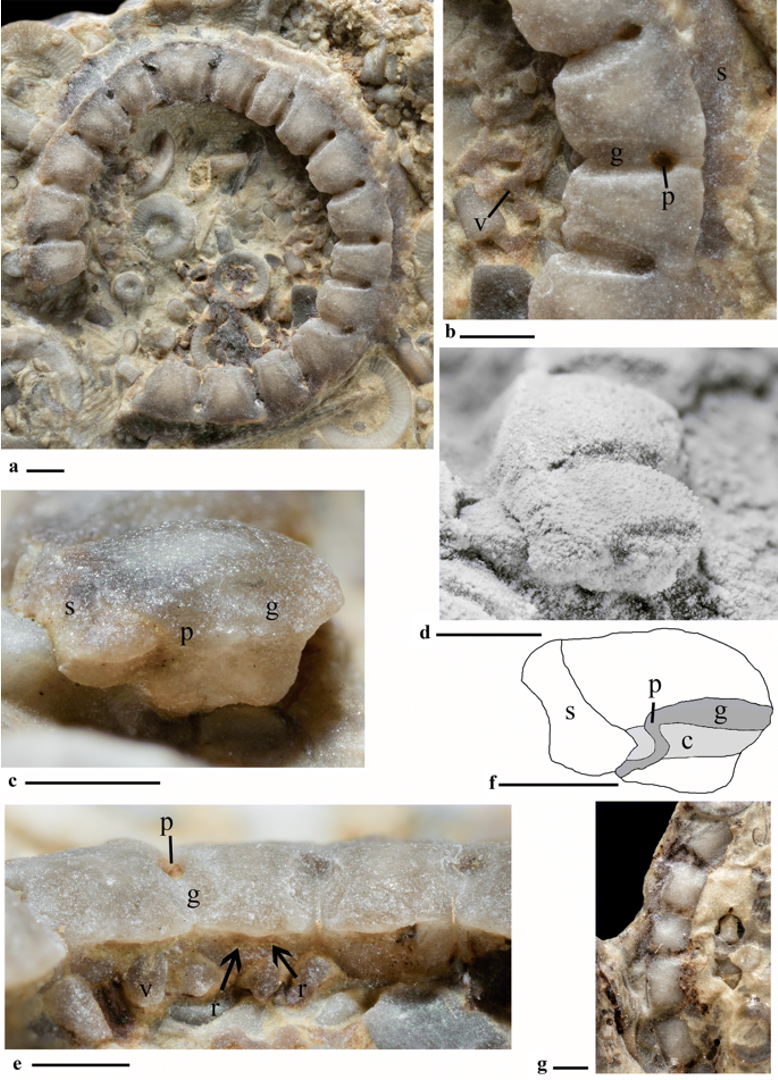
The cyclocystoid "Perforocycloides nathalieae gen. et sp. nov., MPEP335.7. Early Silurian, Upper Llandovery, Jupiter Formation, Cybèle Member. a Dorsal view of entire specimen. b Detail of dorsal marginal ossicle, distal ventral disc plating and peripheral skirt. c Lateral view of marginal ossicle suture. d Same view as Fig. 2c, but coated in ammonium chloride sublimate. e Proximal surface of marginal ossicles. Note prominent crescentic facet area extended well beyond the ventral crest. Arrows point to radial ducts still filled with sediment. f Camera lucida of lateral view of marginal in Fig. 2c, d. Darker grey corresponds to the dorsal sutural groove which leads to a pore and sinusoidal canal that penetrates the contiguous area of the lateral surface (in light grey) and emerges on the distal side of the ventral marginal surface, adjacent to the cupules. g Ventral view of marginal ossicles (top of the crest). c contiguous area of lateral suture of marginal, g groove on distal surface suture between marginal ossicles, p intermarginal sutural pore, r radial ducts, s peripheral skirt, v ventral disc plating." Figure and caption from Ewin et al. (2019) in PalZ (open access); Creative Commons Attribution 4.0 Internationl license.
Edrioasteroidea
The edrioasteroids originated during the Cambrian and went extinct in the Permian. Living primarily attached to hard substrates, including rocks, shell debris, and other organisms (e.g., brachiopods; see the 3D model below), this class was relatively widespread and lived in many environments. In some edrioasteroid groups, the radial symmetry was somewhat modified from the typical pentaradial form. In these groups, the five ambulacra were distributed in a 2-1-2 arrangement, where two sets of two ambulacra branch together initially, before separating into the familiar pentaradial arrangement. This is regarded as the primitive symmetry state of echinoderms and can be seen in the image below, particularly in the two ambulacra branching together in the lower left quadrant.

A fossil edrioasteroid, Carneyella ulrichi, from the Ordovician of Kentucky (USA). Note the 2-1-2 arrangement of the ambulacra branching from the mouth at the center. Image by James St. John; Creative Commons Attribution 2.0 Generic license.
{kind=link}
Edrioasteroids were suspension feeders like many other echinoderm classes. Resembling a flat disc—although life reconstructions commonly indicate an inflated organism—the five ambulacra were arranged radially around a central mouth, with an anus located just below, on the same upward-facing surface. These components rested on top of the main body (i.e., theca), which was composed of calcium carbonate plates (as in most echinoderms). The theca was attached to the hard substrate but presumably had the ability to move slightly.
Fossil specimen of the edrioasteroid Isorophus cincinnatiensis attached to the brachiopod Rafinesquina alternata; from the Ordovician of Ohio (PRI 76783). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Width of brachiopod is 4 cm. Model by Emily Hauf.
Ophiocistoidea
The ophiocistioids partially resemble the extant classes of mobile pentaradial echinoderms and may be ancestral to one or more of these classes. The main body of an ophiocistioid is somewhat similar to the test of an echinoid (i.e., sea urchin), with numerous calcareous plates fitted together, and their five-part jaw is also somewhat similar to the echinoid mouthpiece, Aristotle’s lantern. More similar to the asteroids (i.e., sea stars) and ophiuroids (i.e., brittle stars), the central test is surrounded by a collection of appendages, called podia in this class. The podia were likely used for locomotion, and potentially food capture, much as in modern sea and brittle stars. Interestingly, the podia were also covered in calcium carbonate plates—hence their good preservation in the fossil record—which distinguishes them from the living stars.

Hand drawings of a generalized Ophiocistoidea specimen, with the mouth located centrally (left), and of the species Eucladia woodwardi, viewed from the aborally (top right) and orally (bottom right). Both images are by Charles Schuchert (public domain).
{kind=link}
{kind=link}
Though their general morphology has aspects that resemble asteroids and ophiuroids, ophiocistioid skeletal morphology is more similar to the Echinozoa (i.e., the combined group of echinoids and holothurians), suggesting the class belongs in this grouping. Some researchers have further refined this relationship, such as in the phylogenetic hypothesis above, suggesting that the ophiocistioids are a sister-group of the sea cucumbers (i.e., holothurians). Like many echinoderm classes, the ophiocistioids are first known from the Ordovician and, until recently, were believed to have gone extinct at the end of the Permian. Reich et al. (2018) reported isolated ossicles that were interpreted as a new ophiocistioid from the Late Triassic, extending the record of this group into the Mesozoic.
References and further reading:
Bauer, J. E., C. D. Sumrall, and J. A. Waters. 2017. Hydrospire morphology and implications for blastoid phylogeny. Journal of Paleontology, 91: 847–857.
Deline, B., J. R. Thompson, N. S. Smith, S. Zamora, I. A. Rahman, S. L. Sheffield, W. I. Ausich, T. W. Kammer, and C. D. Sumrall. 2020. Evolution and Development at the Origin of a Phylum. Current Biology, 30: 1672-1679.
Ewin, T. A. M., M. Reich, M. R. Graham, M. E. Cournoyer. 2019. Perforocycloides nathalieae new genus and species, an unusual Silurian cyclocystoid (Echinodermata) from Anticosti Island, Québec, Canada. PalZ, 93: 625-635.
Lefebvre, B., T. E. Guensburg, E. L. Martin, R. Mooi, E. Nardin, M. Nohejlová, et al. 2019. Exceptionally preserved soft parts in fossils from the Lower Ordovician of Morocco clarify stylophoran affinities within basal deuterostomes. Geobios, 52: 27-36.
Nardin, E., B. Lefebvre, O. Fatka, M. Nohejlová, L. Kašička, M. Šinágl, M. 2017. Evolutionary implications of a new transitional blastozoan echinoderm from the middle Cambrian of the Czech Republic. Journal of Paleontology, 91: 672-684.
Nohejlová, M., E. Nardin, O. Fatka, L. Kašička, M. Szabad. 2019. Morphology, palaeoecology and phylogenetic interpretation of the Cambrian echinoderm Vyscystis (Barrandian area, Czech Republic). Journal of Systematic Palaeontology, 17: 1619-1634.
Rahman, I. A., S. E. Stewart, and S. Zamora. 2014. The youngest ctenocystoids from the Upper Ordovician of the United Kingdom and the evolution of the bilateral body plan in echinoderms. Acta Palaeontologica Polonica, 60: 39-48.
Reich, M., T. R. Stegemann, I. M. Hausmann, V. J. Roden, and A Nützel. 2018. The youngest ophiocistioid: a first Palaeozoic‐type echinoderm group representative from the Mesozoic. Palaeontology, 61: 803-811.
Sheffield, S. L., and C. D. Sumrall. 2019. The phylogeny of the Diploporita: a polyphyletic assemblage of blastozoan echinoderms. Journal of Paleontology, 93: 740-752.
Smith, A. B., S. Zamora, and J. J. Alvaro. 2013. The oldest echinoderm faunas from Gondwana show that echinoderm body plan diversification was rapid. Nature Communications, 4: 1-7.
Sprinkle, J., and T. E. Guensburg. 1995. Origin of echinoderms in the Paleozoic Evolutionary Fauna: the role of substrates. Palaios, 437-453.
Sumrall, C. D., and A. G. Wray. 2007. Ontogeny in the fossil record: diversification of body plans and the evolution of “aberrant” symmetry in Paleozoic echinoderms. Paleobiology, 33: 149-163.
Zamora, S., and I. A. Rahman. 2014. Deciphering the early evolution of echinoderms with Cambrian fossils. Palaeontology, 57: 1105-1119.
Zamora, S., B. Deline, J. J. Álvaro, and I. A. Rahman. 2017. The Cambrian Substrate Revolution and the early evolution of attachment in suspension-feeding echinoderms. Earth-Science Reviews, 171: 478-491.
Zamora, S., I. A. Rahman, and A. B. Smith. 2012. Plated Cambrian bilaterians reveal the earliest stages of echinoderm evolution. PLoS One, 7: e38296.
Usage
Unless otherwise indicated, the written and visual content on this page is licensed under a Creative Commons Attribution-NonCommercial-Share Alike 4.0 International License. This page was written by Jansen A. Smith. See captions of individual images for attributions. See original source material for licenses associated with video and/or 3D model content.
