


Section contents:
Feature image: Close-up of a swamp mallow flower (Hibiscus moscheutos) showing stigmas, staminal tube, and petals. Credit: E.J. Hermsen (DEAL).
Introduction
The flower is the reproductive unit of an angiosperm, meaning that it is the location in which the key processes of sexual reproduction (meiosis and fertilization) are carried out. Many flowers have two types of fertile structures, carpels that enclose ovules (immature seeds) and stamens that make pollen. Sterile structures (petals and sepals) that vary in appearance from green and leaf-like to brightly colored may occur below the fertile structures. In a typical flower, both the fertile and sterile structures are interpreted as leaf homologues (modified leaves). The carpels and stamens are thus sporophylls, or fertile leaves (strictly, sporangium-bearing leaves). Furthermore, the whole flower can be considered a type of simple strobilus, or cone, because it is a terminal reproductive unit consisting of a central axis that bears sporophylls.
Flowers vary in the numbers and arrangements of carpels, stamens, petals, and sepals that they possess, as well as in other attributes. Animal-pollinated flowers are often modified in ways that help them to attract and interact with their pollinators, such as having bright colors, patterns, rewards (i.e., nectariferous tissue that produces nectar), and enticing fragrances. Wind-pollinated flowers tend to be inconspicuous, without a lot of unnecessary structures that might get in the way of pollen grains transported on air currents. (Read more about pollination here.) Flowers may have either carpels (ovule-producing structures) or stamens (pollen-producing structures), or both carpels and stamens.
Variations in flower structure are very important in the identification and classification of modern angiosperms. Flowers are ephemeral and often delicate. Thus, except in certain unusual circumstances (for example, amber inclusions, Cretaceous charcoalified floras), flowers tend to be rare in fossil deposits.

Hydrangea (Hydrangea) flowers. Some hydrangeas produce sterile flowers (flowers not capable of reproducing) with large sepals. Left: Japanese climbing hydrangea (Hydrangea anomala). Right: Hydrangea fraxinifolia (Late Eocene, Florissant Formation, Colorado, U.S.A.). Credits: Hydrangea anomala (Dominicus Johannes Bergsma, Wikimedia Commons, CC BY-SA 3.0); Hydrangea fraxinifolia, YPM PB 023931 (Yale Peabody Museum, iDigBio Portal, CC0 1.0/public domain). Images modified from originals.
{kind=link}
Overview of floral structure & terminology
The "ideal" or "stereotypical" flower is made up of four levels, or whorls, of structures attached to an axis called a receptacle. These whorls are as follows, listed from innermost (or uppermost) to outermost (or lowermost):
- Carpels: The megasporophylls (megasporangium-bearing leaves) or "female" parts of a flower. A flower may have one carpel, two or more distinct carpels (i.e., carpels that are not fused to one another), or two or more carpels fused into a single structure. A typical carpel has three regions:
- The stigma, a specialized surface on which pollen grains land and germinate.
- The style, a stalk-like structure that elevates the stigma.
- The ovary, the portion of the carpel that contains the ovules, or the structures that will become seeds following fertilization. In fact, the ovary is what gives angiosperms (Greek, angeion + sperma = vessel + seed) their name. Flowering plants keep their seeds in a container, the fruit. A fruit develops from the ovary of a flower, and the seeds develop from the ovules.
- Stamens: The microsporophylls (microsporangium-bearing leaves) or "male" parts of a flower. A typical stamen has two distinct regions.
- The filament is a thin stalk.
- The anther occurs at the end of the filament. Pollen develops in the anther.
- Petals: The petals are often the "showy" or colorful parts of the flower. These structures are often the parts that make flowers attractive to humans, but also to other animals that may serve as pollinators.
- Sepals: The sepals are often green and leaf-like, although sometimes they may be colorful like the petals.
Note: Female and male are in quotes above because, technically speaking, the carpels and stamens are not themselves female or male. Rather, the carpels enclose the ovules in which the female embryo sacs (egg-producing gametophytes) develop. The male pollen grains (sperm-producing gametophytes) develop in the stamens. For simplicity's sake, however, carpels are sometimes called "female" and stamens are sometimes called "male." Similarly, "female" flowers have only carpels, whereas "male" flowers have only stamens, etc.
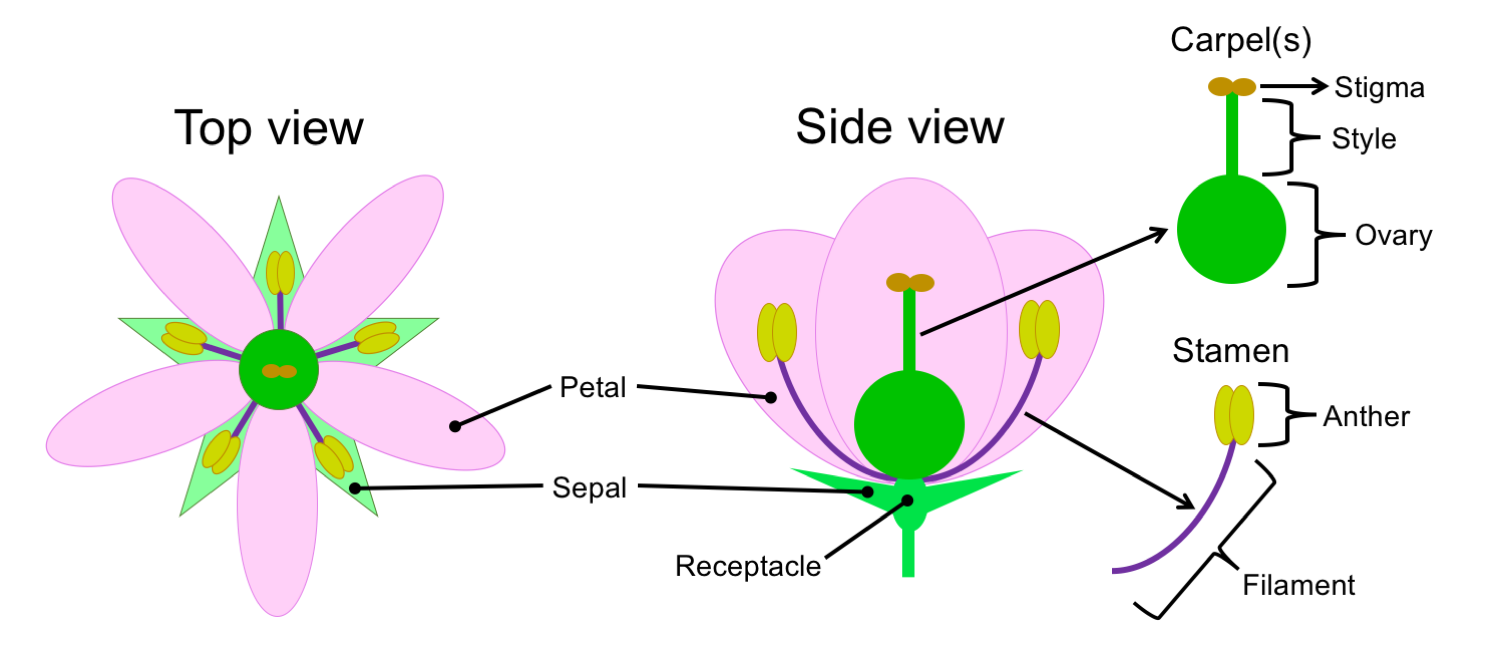
The basic structure of a flower. Credit: E.J. Hermsen (DEAL).
In addition to the terms for the individual structures of a flower, there are terms for the structures as a collective (in other words, all the structures in a single whorl or of a similar type). These are as follows (from uppermost to lowermost whorl):
- Gynoecium (Greek, gynē + oikos = female house): All the carpels in a flower.
- Androecium (Greek, andro + oikos = male house): All the stamens of a flower.
- Corolla: All the petals of a flower.
- Calyx: All the sepals of a flower.
Terms for how the flower is supported:
- Peduncle (solitary flower) or pedicel (one flower in a group of flowers): floral stalk.
- Sessile: lacking a stalk (can refer to flowers, leaves, etc.).
Variation in number/presence of floral whorls
The organization of a flower into four floral whorls is an idealized representation; many flowers do not follow this structure. For example, the floral structures may be helically arranged on the receptacle, as in the stamens and carpels of Magnolia. Sometimes, there may be more than one whorl of the same category of structure, such as stamens.
Another variation is that one or more floral whorls may be missing. For example, a wind-pollinated flower may lack petals, or may have no perianth at all. Functionally, such a flower does not require these structures to attract pollinators. Many times, a flower will be missing either stamens or carpels. Flowers that have both stamens and carpels are bisexual (Latin, bis = two), whereas staminate flowers (flowers having stamens and no carpels) or carpellate flowers (flowers with carpels but no stamens) are unisexual (Latin, unus = one).

Unisexual flowers. Groups of unisexual flowers of gray willow (Salix cinerea). The flowers have a perianth represented only by a nectary (nectary-producing structure). While they lack colorful petals, the flowers are visited by and pollinated by insects. Left: Group of carpellate (pistillate) flowers, each with two carpels. Right: Group of staminate flowers, each with two stamens. Credits: Carpellate inflorescence of Salix cinerea (AnRo00, Wikimedia Commons, CC0 1.0); Staminate inflorescence of Salix cinerea (Krzysztof Ziarnek, Kenraiz, Wikimedia Commons, CC BY-SA 4.0). Images modified from originals.
{kind=link}
{kind=link}
Fusion of structures
Fusion, or lack of fusion, amongst floral parts can radically alter the appearance of a flower. Fusion can occur between structures of the same type (connate structures), structures of different types (adnate structures), both, or neither (distinct or free structures).

Fusion of floral parts. Left: Mount Cook buttercup (Ranunculus lyallii), showing lack of fusion among parts. Center: Field bindweed (Convolvulus arvensis) flower showing corolla tube of connate petals. Right: Primrose (Primula x pubescens) showing epipetalous stamens (Greek, epi = on, over), or stamens that are adnate to the corolla tube (arrow shows one of six visible anthers). Credits: Mount Cook buttercups (Bernard Spragg. NZ, Wikimedia Commons, CC0 1.0); Convolvulus arvensis (AnRo0002, Wikimedia Commons, CC0 1.0); Primula xpubescens (Niepokoój Zbigniew, Wikimedia Commons, CC BY-SA 4.0). Images modified from originals.
.jpg){kind=link}
{kind=link}
{kind=link}
Perianth (sepals, petals & tepals)
The perianth is the collective name for all the sterile structures that form the lowermost (outermost) whorls or helices in a flower. The perianth may be made up of sepals, petals, both sepals and petals, or tepals. Tepals is a term that can be applied when sepals and petals look alike or are not clearly distinguishable from each other. In many flowers, the number of members of each whorl of the perianth is consistent and equal. Thus, a flower may have three sepals and three petals, or four sepals and four petals. There are many variations, however, including flowers with unequal numbers of parts per perianth whorl or no set number of perianth parts.
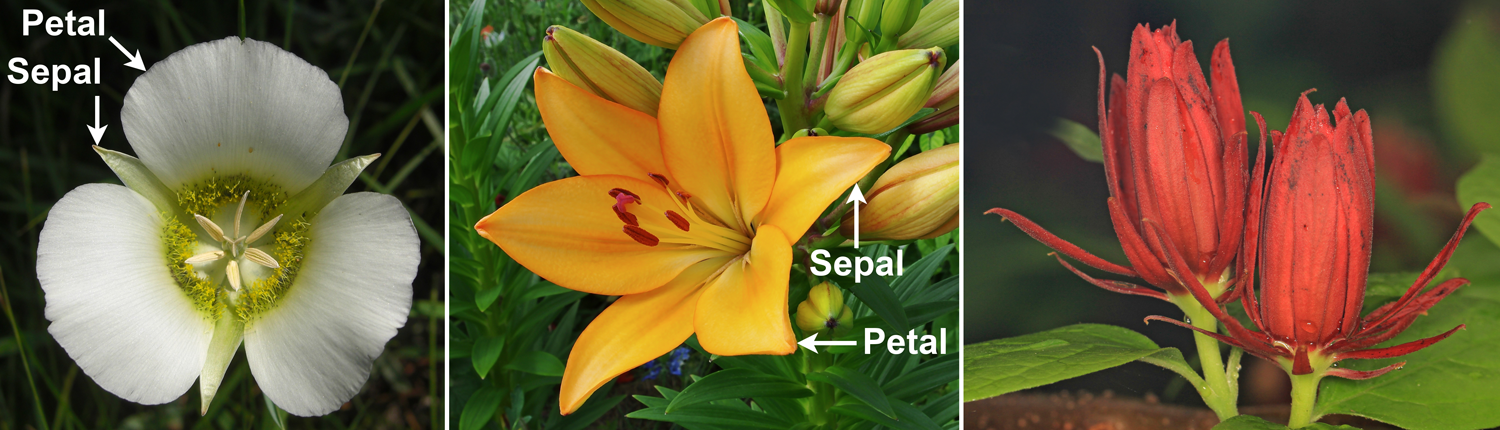
Perianths. Left: Gunnison's mariposa lily (Calochortus gunnisonii) showing 3 sepals that are distinct in appearance from the 3 petals. Center: A cultivated lily (Lilium) with 3 petals and 3 sepals that occur in two whorls; these structures can also be called tepals due to their similarity. Right: Eastern sweetshrub flowers (Calycanthus floridus) with spirally arranged tepals. Credits: Calochortus gunnisonii (Jason Hollinger, flickr, CC BY 2.0); Lilium Lys orange FR 2012 (JLPC, Wikimedia Commons, CC BY-SA 3.0); Calycanthus floridus (Judy Gallagher, Wikimedia Commons, CC BY 2.0). Images modified from originals.
{kind=link}
{kind=link}
Variations in floral symmetry are often especially noticeable in the perianth whorls. Two basic types of floral symmetry are radial and bilateral. A flower with radial symmetry has more than two planes of symmetry, or dividing lines that can be used to cut a flower into symmetrical halves. A flower with bilateral symmetry has only a single plane of symmetry, or a single dividing line that produces symmetrical halves.

Floral symmetry. Left: Zebra-striped huernia (Huernia zebrina), radial symmetry. Right: Japanese ladyslipper (Cypripedium japonicum), bilateral symmetry. Credits: Huernia zebrina subsp. zebrina (SAplants, Wikimedia Commons, CC BY-SA 4.0); Cyprepedium japonicum flower (Alpsdake, Wikimedia Commons, CC BY-SA 4.0). Images modified from originals.
{kind=link}
.JPG){kind=link}
Androecium (stamens)
The androecium includes all the stamens of a flower, which often occur in one or two whorls. The number of stamens in a flower is typically a multiple of the number of perianth members in a single whorl. For example, if a flower has 5 sepals and 5 petals, it might have either 5 stamens or 10 stamens. Stamens are often arranged symmetrically around the center of the flower, in alignment with members of one or both perianth whorls. Stamens can be arranged in other ways. For example, some flowers have many stamens; these may be arranged evenly around the receptacle, or the stamens may occur in fascicles (discrete groups). Others flowers have stamens that are connate (fused) into tubes.

Examples of different types of stamen arrangement in flowers. Left: Flower of encrusted saxifrage (Saxifraga hostii) showing 10 stamens, each aligned with a sepal or a petal. (The sepals are not visible.) The large, yellow ring in the center of the flower is a nectary (nectar-producing structure). Center: Woodland peony (Paeonia obovata) showing many distinct stamens surrounding two carpels. Right: Hibiscus (Hibiscus) showing a tube formed by many stamens; the free ends of the filaments bear the anthers (orange structures). Credits: Saxifraga hostii (Salicyna, Wikimedia Commons, CC BY-SA 4.0); Paeonia japonica (Alpsdake, Wikimedia Commons, CC BY-SA 4.0); Hibiscus (Sander van der Wel, Wikimedia Commons, CC BY-SA 2.0). Images modified from originals.
{kind=link}
.jpg){kind=link}
_Delicate_beauty_(5733278227).jpg){kind=link}
As described above, a typical angiosperm stamen consists of a filament (stalk) that supports an anther (pollen-producing structure). The typical angiosperm anther is made up of four pollen sacs (microsporangia), grouped into two pairs. The microspore mother cells, also called pollen mother cells, differentiate in the pollen sacs and eventually give rise to the pollen grains. The pollen sacs are lined with a specialized tissue called tapetum, which serves as a nutritive (food) tissue for the developing pollen grains. The pollen sacs of the anther open by slits, or sometimes other mechanisms like pores, to release their pollen.
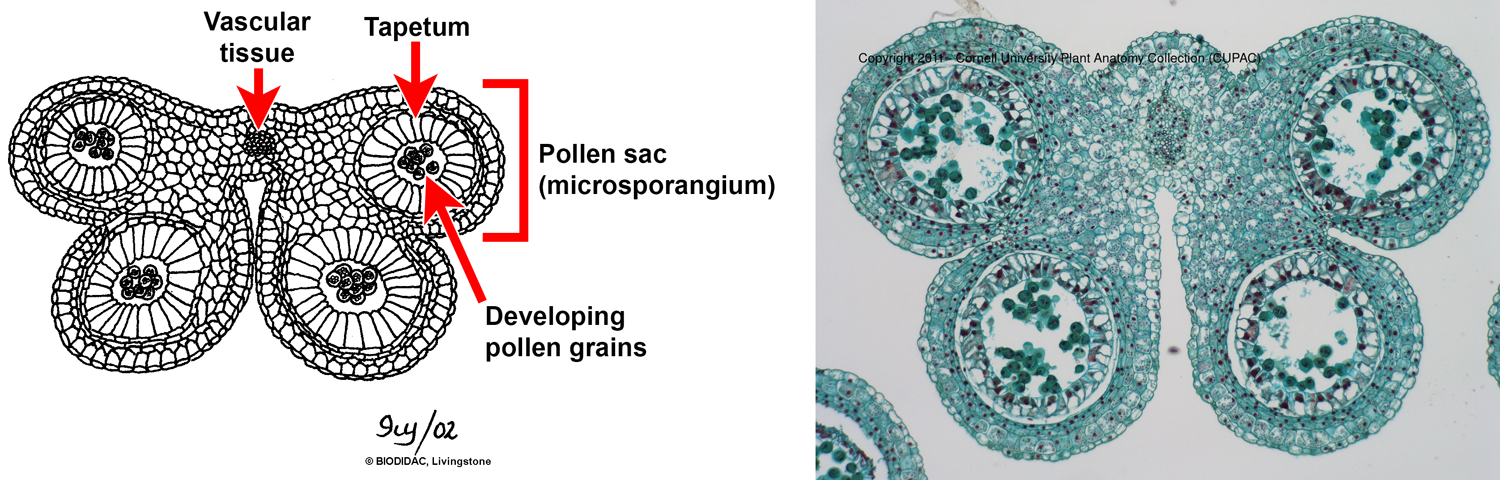
Transverse section of lily (Lilium) anthers showing pollen sacs (microsporangia). Left: Drawing of an anther in transverse section showing four pollen sacs arranged in two pairs. The pollen sacs are lined by the tapetum. Right: Photomicrograph of an anther showing the same structures shown in the labelled drawing. Credits: Transverse section through a lily anther (Ivy Livingstone, BIODIDAC, CC BY-NC 4.0); CUPAC slide 178, image 3 (CUPAC, copyright 2011 Cornell University Plant Anatomy Collection, used with permission). Images modified from originals.
{kind=link}

Dehisced anthers. Left: Detail of a flower (Hippeastrum) showing stamens, some with open (dehisced) anthers and some anthers that have yet to open. Right: Scanning electron micrograph of open Egyptian starcluster (Pentas lanceolata) anthers. Credits: Hippeastrum anthers (Giraffenigel, Wikimedia Commons, CC BY-SA 3.0); SEM of Penta lanceolata anther (Louisa Howard and Charles Daghlian, Dartmouth Electron Microscope Facility, public domain). Images modified from originals.
{kind=link}
Gynoecium (carpels)
The gynoecium includes all the carpels of a flower. A flower may have one to many carpels. If a flower has more than one carpel and the carpels are distinct (not fused to each other), then the gynoecium can be described as apocarpous (Greek, apo = separate). If a flower has more than one carpel and the carpels are connate (fused to one another), then the gynoecium can be described as syncarpous (Greek, syn = together).
Sometimes, the term pistil is used to describe the gynoecium instead of or in addition to the term carpel. One or more carpels make up a pistil. Thus, depending on the structure of the flower, the terms carpel and pistil may or may not be interchangeable. If a flower has a single carpel or an apocarpous gynoecium, the terms are essentially interchangeable. If a flower has a syncarpous gynoecium, then the terms are not interchangeable. Rather, the pistil is the structure made up of all the fused carpels together. It can sometimes be difficult to determine whether a pistil is made up of a single carpel or more than one carpel. In a syncarpous gynoecium, the number of carpels may be indicated by the number of styles, the number of stigma lobes, and/or the number of locules (chambers) in the ovary (see below).

Carpel fusion. Left: Close-up of the center of a southern magnolia (Magnolia grandiflora) flower, showing the apocarpous gynoecium made up of many distinct carpels (pistils) helically arranged on an elongated floral receptacle. Right: Tulip (Tulipa) flower with a 3-carpellate, syncarpous gynoecium (1 pistil of 3 carpels); in this case, the three stigma lobes (indicated by the arrowheads) provide a clue that the flower has three carpels. Credits: Magnolia grandiflora flower (Liné1, Wikimedia Commons, CC BY-SA 2.5); Tulipa flower (Jorgebarrios, Wikimedia Commons, CC BY-SA 3.0). Images modified from originals.
{kind=link}
{kind=link}
The carpel or pistil is divided into at least two, and typically three, distinct regions. These are the stigma, the style, and the ovary. The stigma is the surface on which pollen grains land. The style is a stalk that elevates the stigma, often so that it projects above some or all other floral parts. Not all carpels have a style.
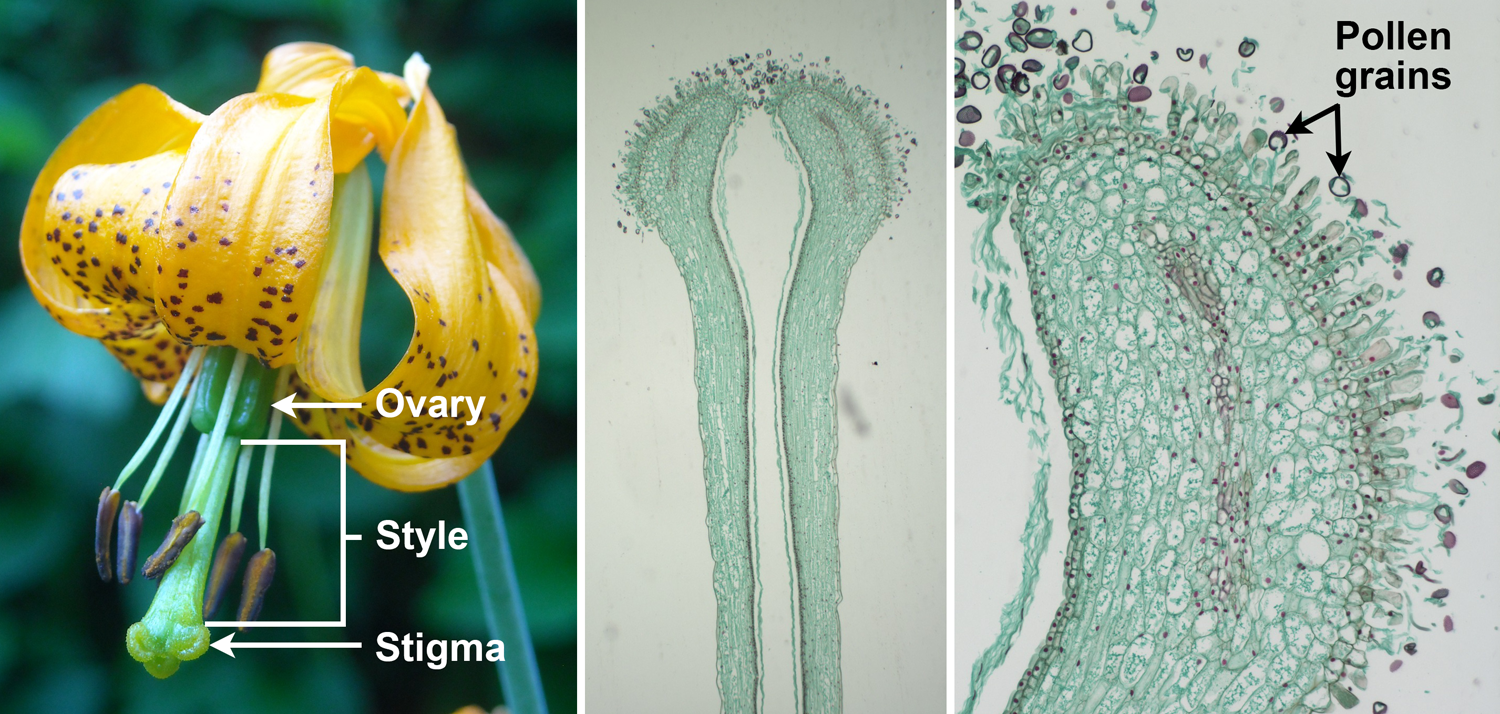
Carpel regions. Left: Tiger lily (Lilium columbianum) flower with pistil of three carpels, showing stigma, style, and ovary regions. Center: Longitudinal section of the distal portion of a lily (Lilium) style capped by stigmatic tissue. Right: Longitudinal section of a portion of a lily (Lilium) stigma showing the papillate surface upon which pollen grains have landed. Credits: Tiger lily (NPS/Chris Roundtree, Wikimedia Commons, Public Domain); CUPAC slide 191, image 1 (CUPAC, copyright 2011 Cornell University Plant Anatomy Collection, used with permission); CUPAC slide 191, image 8 (CUPAC, copyright 2011 Cornell University Plant Anatomy Collection, used with permission). Images modified from originals.
.jpg){kind=link}
{kind=link}
{kind=link}
The ovules are contained within the region of the carpel(s) known as the ovary, which is at the base of and often wider than the style. The ovary may have a single locule (chamber), or may be divided into two or more locules internally. If the ovary is subdivided into two or more locules, the locules are separated by partitions known as septa. In an ovary made up of more than one fused carpel, the locule number may be the same as the carpel number. It should be noted, however, that sometimes fused carpels are not separated by septa and the locule number is one. In other cases, there may be more locules than carpels.
Ovules may be attached at the periphery of the ovary wall, near the junctions of the septa, at the top of the ovary, at the base of the ovary, or on a central column. These different arrangements of the ovules are called placentation. In angiosperms, the placenta is the region where the ovules attach to the ovary wall.
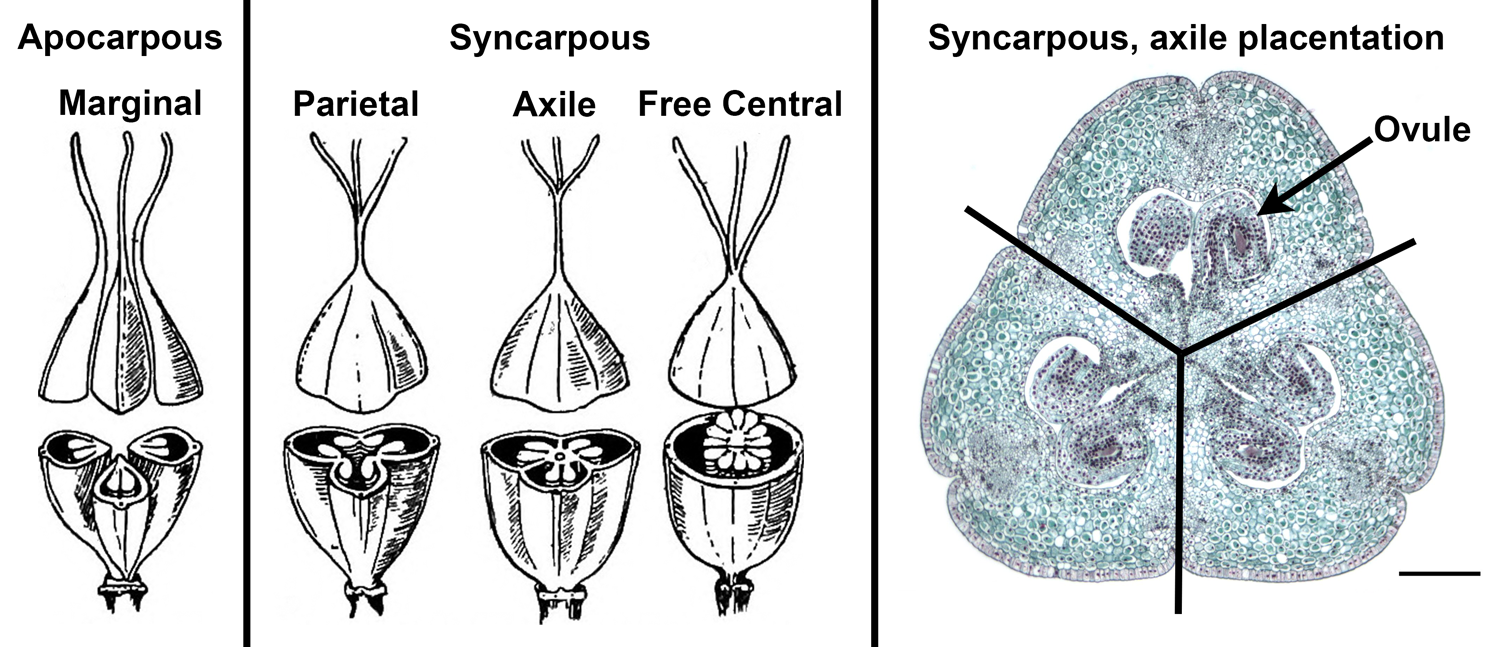
Internal organization of the ovary. Left: Illustrations showing examples of variation in carpel fusion, placentation, and locule number in a gynoecium with three carpels. The apocarpous gynoecium has three distinct (unfused) carpels with marginal placentation. Three types of syncarpous placentation are shown in the syncarpous gynoecia. In parietal placentation, there is one locule and ovules are attached to the ovary wall where the edges of the carpels are fused. In axile placentation, the carpels are separated by septa, and the ovules are attached in the center of the ovary. In free central placentation, the ovules are attached to a central column. Right: Transverse section of a lily (Lilium) ovary that has three fused carpels. The black lines, which are aligned with the septa, separate the carpels. Note that the placentation is axile, and six ovules (two per carpel) are attached near the center of the ovary. Credits: Image from page 79 of Elementary Botany, 1898, author Percy Groom (digitized by Internet Archive, flickr, no known copyright restrictions); Lilium ovary (Jon Houseman and Matthew Ford, Wikimedia Commons, CC BY-SA 4.0). Images modified from originals.
{kind=link}
Ovary position & insertion of parts
The carpels often do not appear to be the uppermost whorl of floral parts. In fact, ovary position and insertion of parts are considered important characteristics in flowering plant identification and classification. Ovary position can also be important in understanding fruit structure. There are two basic terms to describe ovary position and three basic terms to describe variation in the insertion of floral parts:
- Hypogynous (Greek, hypo + gynē = below woman): The perianth and stamens are attached below the ovary. The ovary in this type of flower is considered superior, or above the other floral parts.
- Perigynous (Greek, peri + gynē = around woman): The perianth and stamens are attached to the rim of a hypanthium, or floral cup. The hypanthium surrounds but is not fused to the ovary wall. The ovary is still considered to be superior because it is attached above the hypanthium.
- Epigynous (Greek, epi + gynē = above woman): The perianth and stamens appear to be attached at the top of the ovary. The ovary is considered inferior, or below the other floral parts. In this type of flower, the ovary is interpreted as being sunken into the floral receptacle or surrounded by the bases of the other floral whorls, which are fused to the outer ovary wall.
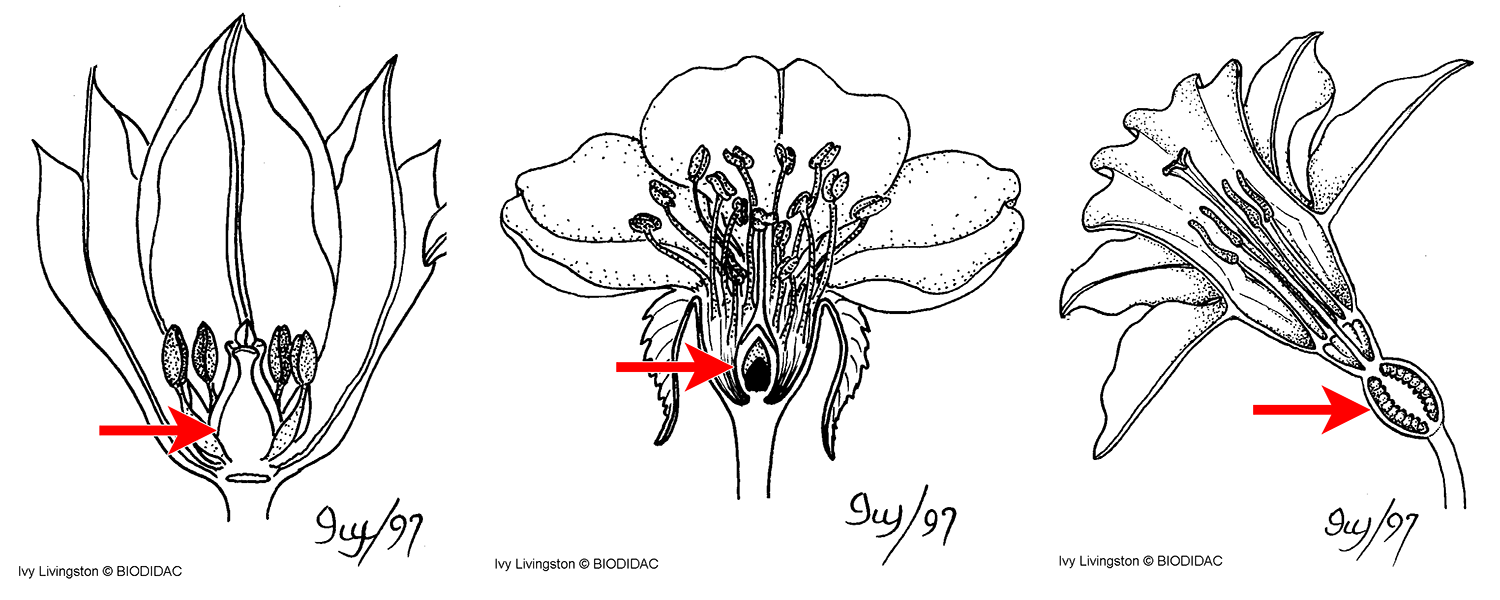
Ovary position and insertion of parts. On each drawing, the location of the ovary is indicated by a red arrow. Left: A hypogynous flower (superior ovary). Center: A perigynous flower (superior ovary surrounded by free hypanthium). Right: An epigynous flower (inferior ovary). Credits: Hypogynous flower, perigynous flower, and epigynous flower (Ivy Livingston, BIODIDAC, CC BY-NC 4.0). Images modified from originals.
Beyond the four whorls: other structures
Flowers may have additional structures that do not simply fall into the category of sepal, petal, stamen, or carpel. Some examples include the nectary, the staminode, and the corona.
Many flowers have a nectary or nectaries made up of specialized tissue that exudes nectar. Sometimes, nectar-producing tissue may be inconspicuous. At other times, it may form an obvious floral structure or structures, often a conspicuous ring or "doughnut" inside the stamens that encircles the pistil or ovary.
Staminodes are sterile stamens, sometimes highly modified. These may be functional, typically in pollination, or may have no particular function.
A corona is a structure or series of structures that occurs between the corolla (petals) and androecium (stamens) in a flower. The corona has evolved independently in several different groups of flowers, and is variable in form. For example, in the daffodil (Narcissus), the corona is a petal-like tube.

Nectaries, staminodes, and coronas. Left: Wilga (Geijera parviflora) flower showing large, yellow nectary. Center: Palmer's penstemon (Penstemon palmeri), showing the prominent fuzzy staminode that gives the beardtongues (Penstemon) their name (four anthers are visible inside the mouth of the corolla). Right: Daffodil (Narcissus pseudonarcissus) showing the tube-like corona. Credits: Wilga-Havilah3 (Poyt448 Peter Woodard, Wikimedia Commons, CC0 1.0); Penstemon palmeri 3 (Stan Shebs, Wikimedia Commons, CC BY-SA 3.0); Narcissus pseudonarcissus (James Petts, Wikimedia Commons, CC BY-SA 2.0). Images modified from originals.
{kind=link}
{kind=link}
.jpg){kind=link}
Inflorescences
Although flowers can be solitary (borne alone on a floral stalk), often they are grouped into aggregations called inflorescences. Inflorescence structure varies widely amongst angiosperms, and can, in some cases, be characteristic of particular groups of plants. The common stalk of an inflorescence is a penduncle. The main axis of an inflorescence (in other words, the axis from which flowers or flower-bearing branches emerge) is a rachis.
Broadly speaking, inflorescences can be divided into two types:
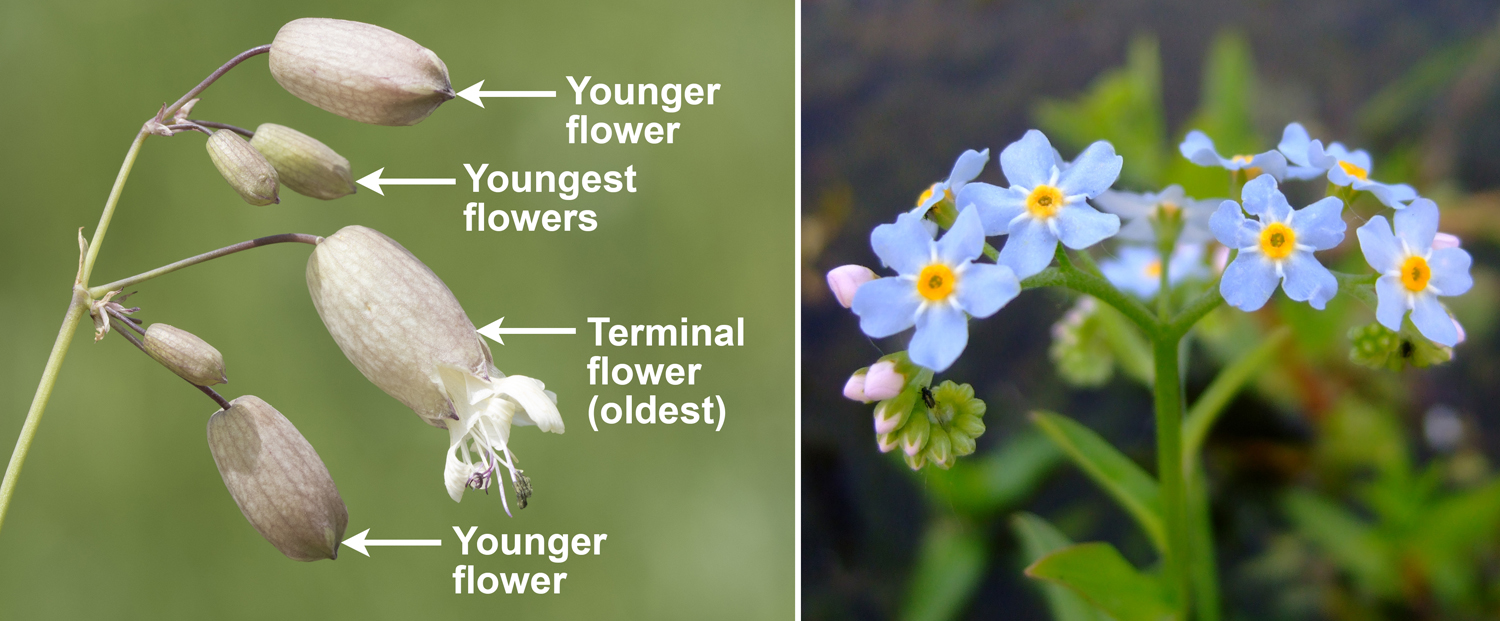
- Indeterminate: In this category of inflorescence, there is no terminal flower on the tip of the inflorescence axis. The least mature (youngest) flower is nearest the apex of the inflorescence. Examples include the panicle, raceme, and spike.
- Determinate: This category of inflorescence is made up of multiple branches, each ending in a flower. When a flower develops at the end of a branch, it prevents further growth of that branch (this is why this type of inflorescence is called determinate; in botany, determinate = limited growth). Inflorescence growth can be continued by one or more lateral (side) branches, each of which will itself end in a flower. Determinate inflorescences include different types of cymes. (To learn more about the branching structure of cymes, read "Sympodial growth of angiosperm inflorescences" on the Branching page in the Introduction to Vascular Plant Structure section.)

Indeterminate inflorescences. Left: Inflorescence (raceme) of foxglove (Digitalis purpurea); note that oldest flowers are at the bottom and the youngest at the top. Center: Inflorescence (spike) of white turtlehead (Chelone glabra); note that the youngest flowers are near the top of the inflorescence. Right: Cow parsnip (Heracleum sphondylium) inflorescence (compound umbel); note that the youngest flowers are in the center of each group of flowers. Credits: Digitalis purpurea (Norbert Nagel, Wikimedia Commons, CC BY-SA 3.0); Chelone glabra 05 (Michael Wolf, Wikimedia Commons, CC BY-SA 3.0); Heracleum sphondylium (Rosser1954, Wikimedia Commons, CC BY-SA 4.0). Images modified from originals.
{kind=link}
{kind=link}
,_Lainshaw,_East_Ayrshire.jpg){kind=link}
{kind=link}
{kind=link}
In some groups, the inflorescence has become so modified that an entire group of flowers resembles a single flower. Examples of plants with this type of inflorescence include some dogwoods (Cornus) and members of the aster family (Asteraceae, like the sunflower below). Some members of the spurge family (Euphorbiaceae) also show this tendency. In poinsettia (Euphorbia pulcherrima), the "flower" is actually a double inflorescence. It is made up a cluster of cyathia (tiny, highly modified inflorescences) surrounded by petal-like bracts!

Flower-like inflorescences. Left: Bunchberry (Cornus suesica), group of flowers surrounded by four white bracts (modified leaves). Right: Sunflower (Helianthus annuus), showing central disk flowers surrounded by ray flowers with large petals. Credits: Cornus suescia (fi:Käyttäjä:kompak, Wikimedia Commons, CC BY-SA 3.0); Helianthus-VallejoSunflower (Jeffreyrea, Wikimedia Commons, CC BY 3.0). Images modified from originals.
{kind=link}
{kind=link}
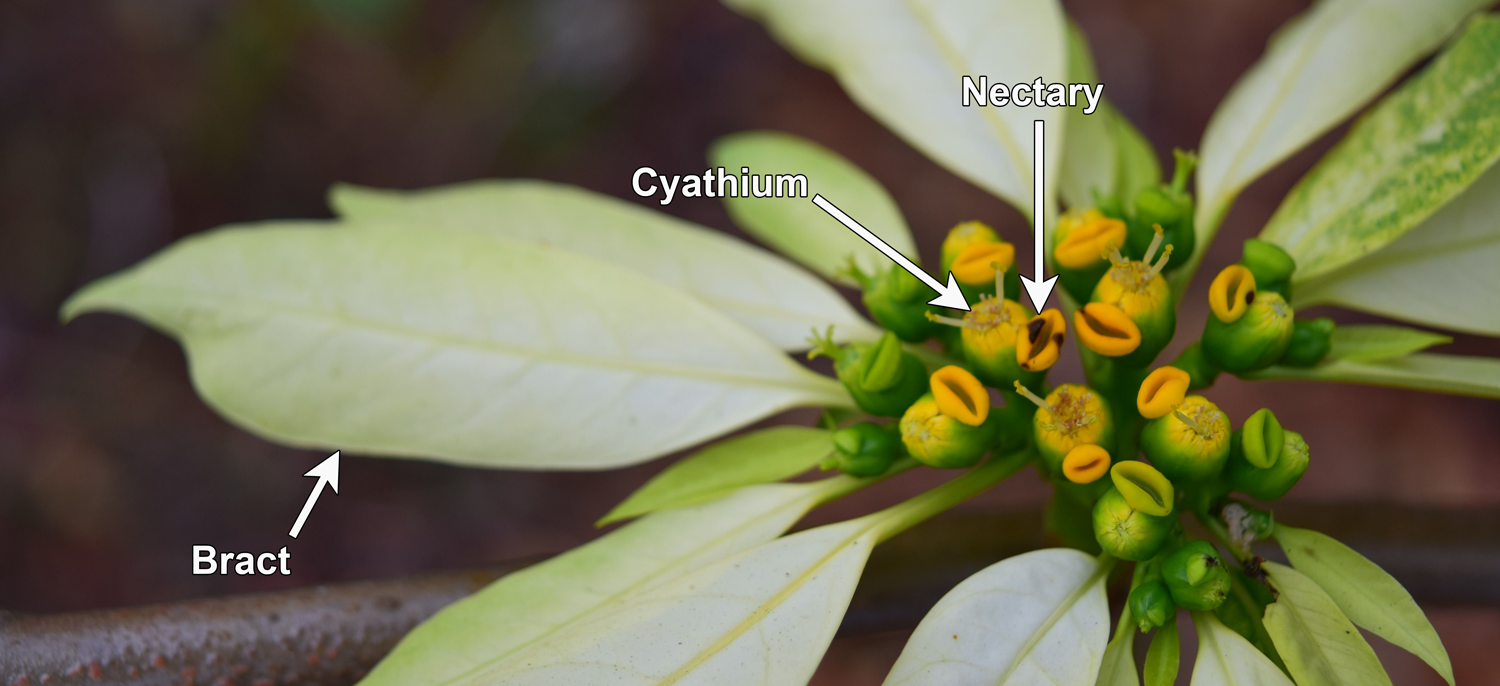
Flower-like inflorescence of poinsettia. The "flower" of poinsettia (Euphorbia pulcherrima) is actually a group of tiny inflorescences surrounded by enlarged bracts, which are often red (although in this image, they are white). Credits: Poinsettia (Arul Raj C, Wikimedia Commons, CC BY-SA 4.0). Image modified from original.
{kind=link}
Selected references & further reading
Note: Free full text is made available by the publisher for items marked with a green star.
Academic articles & book chapters
Rudall, P.J. 2013. Identifying key features in the origin and early diversification of angiosperms. In: B.A. Ambrose and M. Purugganan, eds. The Evolution of Plant Form. Annual Plant Reviews 45: 163-188. https://doi.org/10.1002/9781118305881.ch6
* Walker-Larsen, J., and L.D. Harder. 2011. The evolution of staminodes in angiosperms: patterns of stamen reduction, loss, and functional re-invention. American Journal of Botany 87: 1367-1384. https://doi.org/10/2307/2656866
* Waters, M.T., A.M.M. Tiley, E.M. Kramer, A.W. Meerow, J.A. Langdale, and R.W. Scotland. 2013. The corona of the daffodil Narcissus bulbocodium shares stamen-like identity and is distinct from the orthodox floral whorls. The Plant Journal 74: 615-625. https://doi.org/10.1111/tpj.12150
Books & textbooks
Evert, R.F., and S.E. Eichhorn. 2013. Raven Biology of Plants, 8th ed. W.H. Freeman and Co., New York, New York.
Harris, J.G., and M.W. Harris. 2001. Plant Identification Terminology, an Illustrated Glossary, 2nd ed. Spring Lake Publishing, Spring Lake, Utah.
Meyer, H.W. 2003. The fossils of Florissant. Smithsonian Books, Washington and London.
Simpson, M.G. 2010. Plant Systematics, 2nd ed. Academic Press, Burlington, Massachusetts.
Websites
* In Defense of Plants: The sterile flowers of hydrangea. June 28, 2017. http://www.indefenseofplants.com/blog/2017/6/20/the-sterile-flowers-of-hydrangea
* Flora of North America Editorial Committee, eds. 1993 onwards. Flora of North America North of Mexico. 20+ volumes. New York and Oxford. http://www.fna.org
* New York Botanical Garden Steere Herbarium: Glossary for Vascular Plants. http://sweetgum.nybg.org/science/glossary/
* Stevens, P.F. 2001 onwards. Angiosperm phylogeny website [continuously updated]. http://www.mobot.org/MOBOT/research/APweb
Content usage
Usage of text and images created for DEAL: Text on this page was written by Elizabeth J. Hermsen. Original written content created by E.J. Hermsen for the Digital Encyclopedia of Ancient Life that appears on this page is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Original images and diagrams created by E.J. Hermsen are also licensed under Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
Content sourced from other websites: Attribution, source webpage, and licensing information or terms of use are indicated for images sourced from other websites in the figure caption below the relevant image. See original sources for further details. Attribution and source webpage are indicated for embedded videos. See original sources for terms of use. Reproduction of an image or video on this page does not imply endorsement by the author, creator, source website, publisher, and/or copyright holder.
Adapted images. Images that have been adapted or remixed for DEAL (e.g., labelled images, multipanel figures) are governed by the terms of the original image license(s) covering attribution, general reuse, and commercial reuse. DEAL places no further restrictions above or beyond those of the original creator(s) and/or copyright holder(s) on adapted images, although we ask that you credit DEAL if reusing an adapted image from the DEAL website. Please note that some DEAL figures may only be reused with permission of the creator(s) or copyright holder(s) of the original images. Consult the individual image credits for further details.
Page last updated 27 January 2021.