

Chapter contents:
Evolution and the Fossil Record
– 1. Natural selection
– 2. Species and species concepts
– 3. Speciation
– 4. Punctuated equilibria and stasis ←
–– 4.1 Videos about punctuated equilibrium and stasis
– 5. Macroevolution
–– 5.1 Hierarchies
–– 5.2 Species selection
–– 5.3 Abiotic vs. biotic causes of macroevolution
–– 5.4 Evolutionary radiations
So far in this chapter, you have learned what species are (Sec. 2), how natural selection has honed species to make them well adapted to the environments in which they live (Sec. 1), and how the process of speciation operates (Sec. 3). Now we want to consider what happens to species after they originate. When do they undergo the most morphological change: during or after speciation? How long do species persist after they originate? Answering these fundamental questions is critically important if we want to understand the major evolutionary trends that are apparent from the history of life (including modern biodiversity) and discern the processes that may have produced these patterns.
You learned in the last section that biologists and paleontologists think that allopatric speciation (driven by the geographic isolation of populations) is the process that is (and has been) the most important for the generation of new species. Presuming that this is true, what should we expect to observe when we study the history of species through deep time?
Charles Darwin published only one figure in On the Origin of Species (shown below), and it was very influential. It shows a series of hypothetical evolutionary lineages that are slowly changing over time. Some of these lineages persist; others go extinct.

The only figure from Darwin's (1859) work On the Origin of Species. The original caption simply states, "Diagram of divergence of taxa" (public domain).
This view of gradual, continuous evolutionary change permeated biology and paleontology for many decades after Darwin. The diagram below—redrawn from a similar illustration published in a 1952 paleontology textbook—shows a pattern of hypothetical evolution similar to that presented by Darwin: a particular feature gradually changes over time. For example, this figure might show the thickness of a clam's shell or the length of a bird's beak gradually increasing within a lineage over thousands of years (with a certain amount of variation present during each time interval, represented by the bell curves).
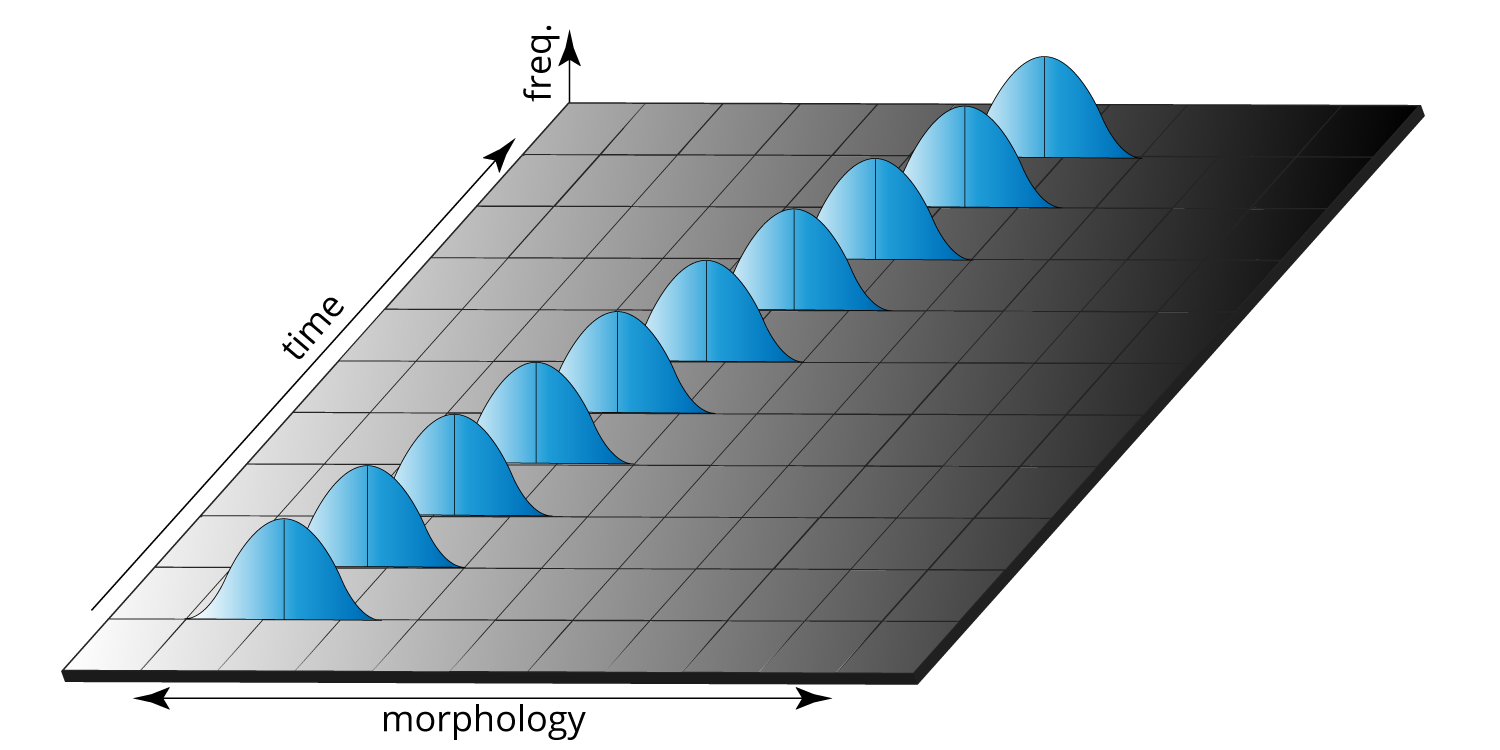
Gradual evolutionary change in a feature over time. The bell curves represent natural variation in that feature during any one particular time interval. Image by Jonathan R. Hendricks redrawn from figure 1-14 in Moore et al. (1952) (it was also republished by Eldredge and Gould, 1972).
For many years, most paleontologists accepted the Darwinian view of evolution that suggested that we should see example after example in the fossil record of gradual, continuous change over time. Supposing this view is correct, what would a perfect fossil record look like? In short, it should show a continuous sequence of ancestral and descendant species, with every intermediate form in between (also called anagenesis).
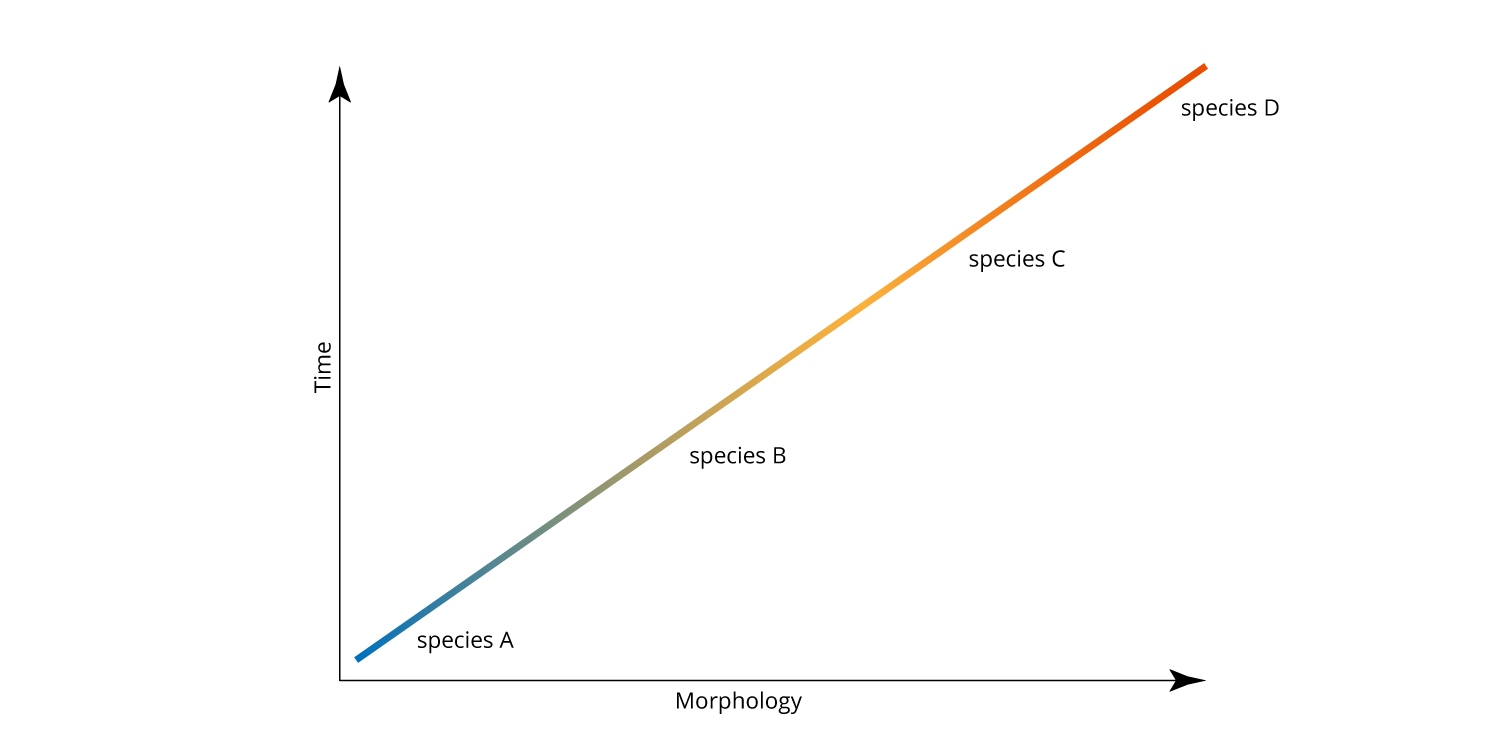
A hypothetical example of anagenesis. Evolutionary change is continuous and ancestral and descendant species are connected by every intermediate form in between. Image by Jonathan R. Hendricks.
The expectation of paleontologists was that they should be able to walk up to a fossiliferous outcropping of rock and observe fossils slowly and seamlessly transforming between successive layers of rock, making it essentially impossible to delineate where one species ended and the next began.
The fossil record, of course, is very far from complete and the expectation presented above was rarely encountered in nature (learn more about why this is the case in the Nature of the Fossil Record chapter). Darwin was very well aware of the incompleteness of the fossil record (in fact, he devoted two chapters to the problem: "On the Imperfection of the Geological Record" and "On the Geological Succession of Organic Beings"). In particular, Darwin knew that most of the intermediate forms that he thought must have existed in the past were unlikely to have been fossilized, much less found.
Following over 160 years of subsequent research on Earth's past, we now have a much better understanding of the fossil record and know that it is not nearly as poor as Darwin suggested (relatively little was known about the fossil record during Darwin's time). Even so, the model of slow, gradual evolutionary change that Darwin promoted remained influential to paleontologists for the next 100+ years. Paleontologists accepted the model of gradual evolution illustrated above, but realized that it would be rare to observe in nature, as the imperfection of the fossil record leaves behind many breaks that make it difficult to directly connect ancestral species and their descendants.
Everything changed, however, in the early 1970s.
Punctuated equilibria
In a classic research paper published in 1972, paleontologists Niles Eldredge and Stephen Jay Gould cast significant doubt on this longstanding assumption that the fossil record should show slow, gradual evolution of one species into the next.
The fundamental question that they asked was, "If allopatric speciation is true, what should the fossil record look like?" Recall from the previous section that allopatric speciation is the model of evolution that emphasizes the geographic isolation of populations—which can cause rapid evolutionary change—as being the principal driver of speciation.
First, they argued that because allopatric speciation happens very quickly on geological time scales, one should not expect to see the gradual change of one species into another in the fossil record. Instead, they stated that new species should most often appear suddenly in the fossil record, reflecting the expanding range and population size of the new daughter species after it migrates back into the range of its parent species. (By their very nature, it is unlikely in most cases that small, geographically isolated populations will be sampled during the geologically short time interval of speciation; see additional discussion below.) Further, newly evolved species should be morphologically distinct relative to their ancestors once they become abundant enough to appear in the fossil record because most morphological change occurs during the interval of geographic isolation.
Second, they argued that after species originate, their morphologies should remain more-or-less constant over time. That is, species should show net morphological stasis throughout their history, such that the oldest and youngest representatives of a species cannot be distinguished from one another.
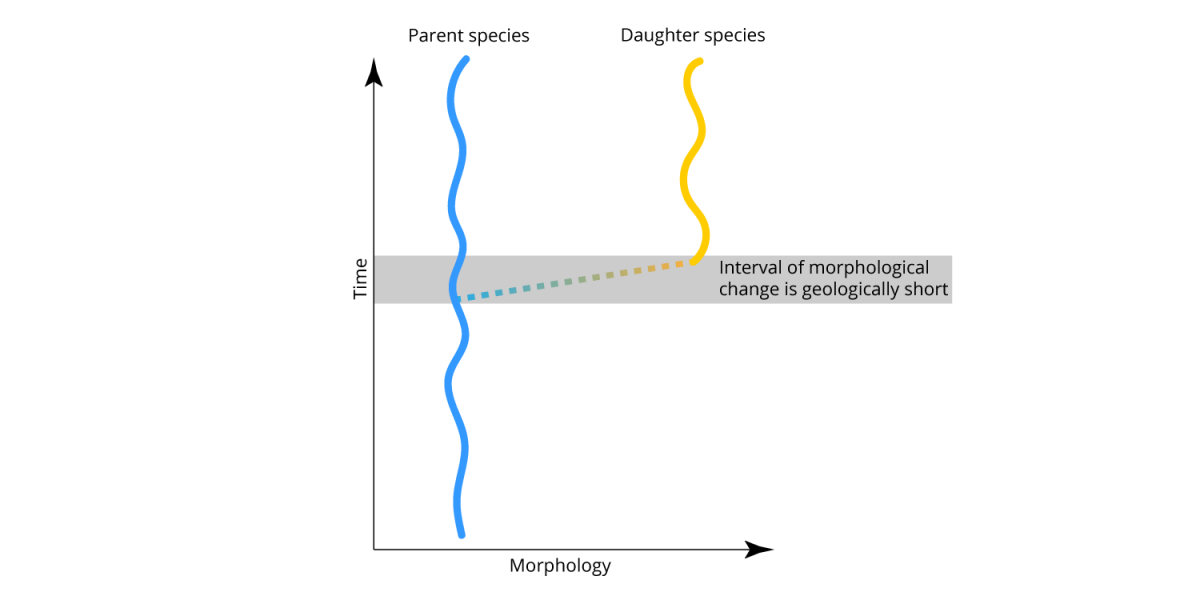
Eldredge and Gould characterized their model as punctuated equilibria. "Punctuated" referred to the sudden appearance of morphologically distinct new species in the fossil record. "Equilibria" referred to the stasis that species exhibit following speciation. Importantly, Eldredge and Gould did not invoke any evolutionary processes that were not already well accepted by evolutionary biologists at the time. Essentially, punctuated equilibria is the theory that incorporated allopatric speciation into the study of species in the fossil record. A generalized model of punctuated equilibrium is shown below. Compare it with the model of slow, steady anagenesis shown above.
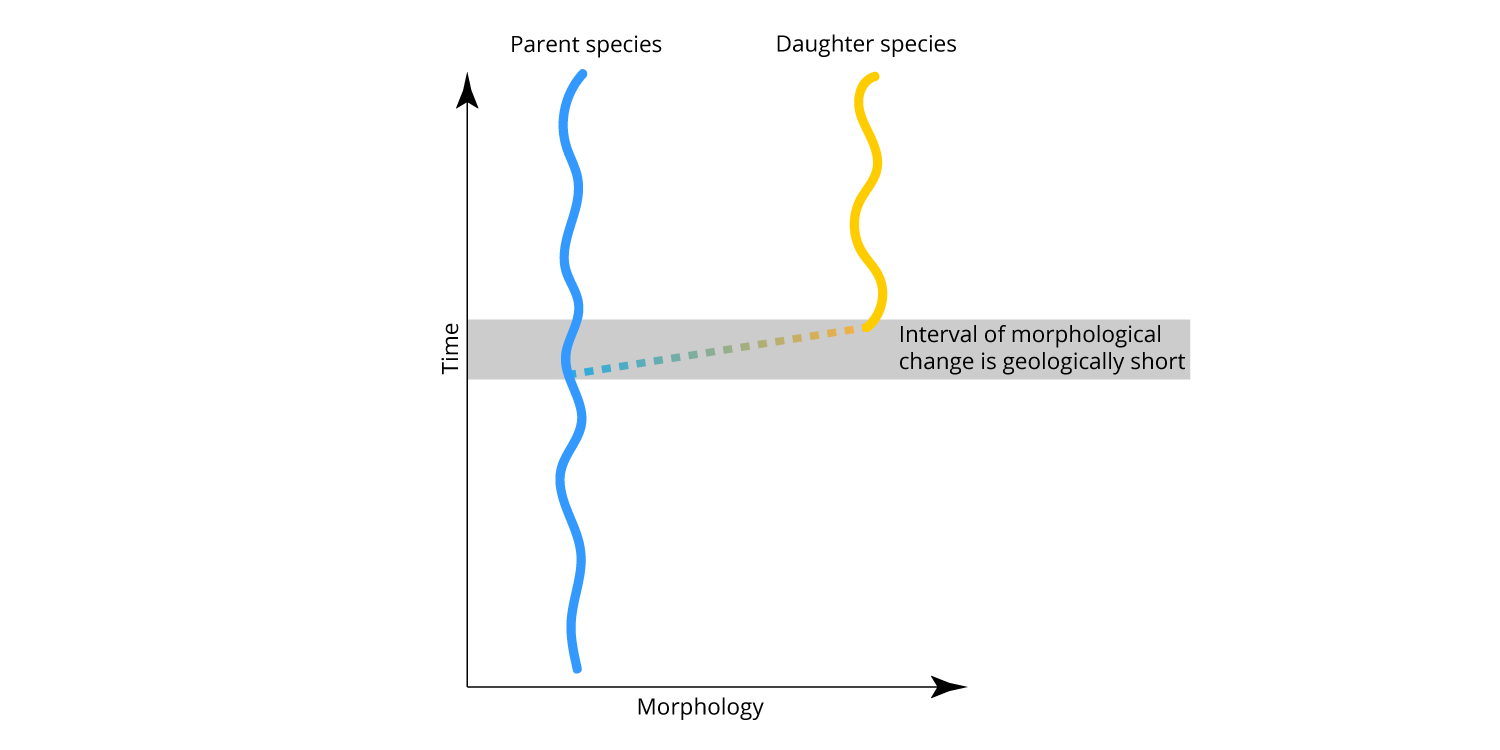
Eldredge and Gould's model of punctuated equilibrium. Note that the interval of speciation (and morphological change) is geologically short (or, punctuated). After species originate, they do not change significantly (i.e., equilibrium or stasis). Image by Jonathan R. Hendricks.
Punctuated equilibria in trilobites and land snails
In their original paper on punctuated equilibria, Eldredge and Gould (1972) provided examples of the pattern in trilobites and land snails.
Eldredge focused on a group of Devonian trilobites, including Phacops rana (later moved to the genus Eldredgeops in honor of Niles Eldredge) and its close relatives, that lived between about 380 and 375 million years ago when a shallow seaway covered much of eastern North America.

Eldredgeops (or, Phacops) rana from the Devonian Ludlowville Formation of Genesee County, New York (PRI 49811). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. A 3D interactive model of this specimen is immediately below. Image by Jonathan R. Hendricks.
Interactive 3D model of the Devonian trilobite Eldredgeops (or, Phacops) rana from the Devonian Ludlowville Formation of Genesee County, New York (PRI 49811). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Trilobite specimen is approximately 3 cm in length. Model by Emily Hauf. A photograph of this specimen is shown above.
Eldredge discovered two principal evolutionary patterns in the trilobites. The first was that new species of Phacops evolved rapidly, effectively in a geological instant—about 5,000 to 50,000 years (more on this below)—in geographically isolated populations. This exactly matched the predictions of allopatric speciation. The second pattern he observed was that once new species appeared, they changed little if at all morphologically. Moreover, these species then persisted for several million years before going extinct and during this entire time they possessed a relatively similar anatomy.
This is not to say that they didn’t change at all. There were some oscillations in morphology, so species weren’t completely invariant, but the net sum of any changes usually tended to cancel out, leading to no net change. This is represented by the wiggly vertical lines in the illustration of punctuated equilibrium shown above.
Gould found very similar patterns in his studies that focused on very different organisms from a very different time period: fossil land snails from the genus Poecilozonites that lived along the beaches of Bermuda a few hundred thousand years ago during the Pleistocene.

Poecilozonites bermudensis, one of the land snail species studied by Stephen Jay Gould in the original formulation of punctuated equilibria (Eldredge and Gould, 1972). Specimen is from the Pleistocene of Bermuda and is from the collections of the Paleontological Research Institution, Ithaca, New York (PRI 76831). Image by Jonathan R. Hendricks.
Rampant stasis in the fossil record
Eldredge and Gould cogently argued that stasis was an important evolutionary phenomenon that had previously largely been ignored by the scientific community. Remember from above that Darwin predicted that the fossil record would show a pattern of anagenesis (or, in the words of Eldredge and Gould, "an insensibly graded fossil sequence" composed of "all transitional forms linking an ancestor with its presumed descendant" (Eldredge and Gould, 1972, p. 87).
Intrepid paleontologists immediately set out to test the prediction of Eldredge and Gould that the fossil record predominantly shows a pattern of morphological stasis, not a pattern of anagenesis. They scaled cliff faces, collected innumerable fossils across strata of different ages, and carefully measured the morphologies of the fossils back in the lab. An example of some of the kinds of measurements that paleontologists captured to document morphological changes are shown in the figures below.

Example of a measurement being collected from a specimen of the brachiopod Mediospirifer audaculus using a digital calipers. Specimen is from the Middle Devonian Moscow Formation of Livingston County, New York (PRI 76830). Image by Jonathan R. Hendricks.

Examples of morphological measurements (red lines) collected from specimens of the brachiopod Mediospirifer audaculus as part of a study of stasis by Bruce Lieberman et al. (1995). Specimen shown is from the Middle Devonian Moscow Formation of Livingston County, New York (PRI 76830). Image by Jonathan R. Hendricks.
Interactive 3D model of Mediospirifer audaculus from the Middle Devonian Moscow Formation of Livingston County, New York (PRI 70767). Specimen is from the collections of the Paleontological Research Institution, Ithaca, New York. Longest dimension of specimen is approximately 5 cm. Model by Emily Hauf.
What did the paleontologists find after they explored the fossil record for evidence of stasis? For the most part they found example after example of morphological stasis, but very few examples of anagenesis. It appeared that morphological stasis was the dominant feature of species in the fossil record.
Why does stasis happen? There are likely several mechanisms that ensure that overall species morphology undergoes little net change, and determining these various mechanisms of stasis is an area of active research. However, one possible explanation is that most species are simply made up of individual organisms that are good enough to get the job done, and thus will persist, unless the pace or amount of environmental change becomes too rapid or profound. In essence, once newly established species become sufficiently abundant to be found in the fossil record they are well enough adapted to persist a long time.
Another pattern germane to the theory of punctuated equilibria is that different clades seem to have different average species durations. For instance, ballpark estimates for the average duration of a species of marine invertebrate such as a trilobite or clam is 5–10 million years. By contrast, species of mammals persist for an average of about 1-2 million years. In addition, groups with shorter species durations, and thus higher likelihoods of going extinct, are almost certainly also evolving more rapidly.
Cladogenesis and sampling speciation
Unlike the Darwinian model of anagenesis (see figure above), punctuated equilibrium emphasized that evolution at the species level involved lineage splitting, or what is termed cladogenesis.
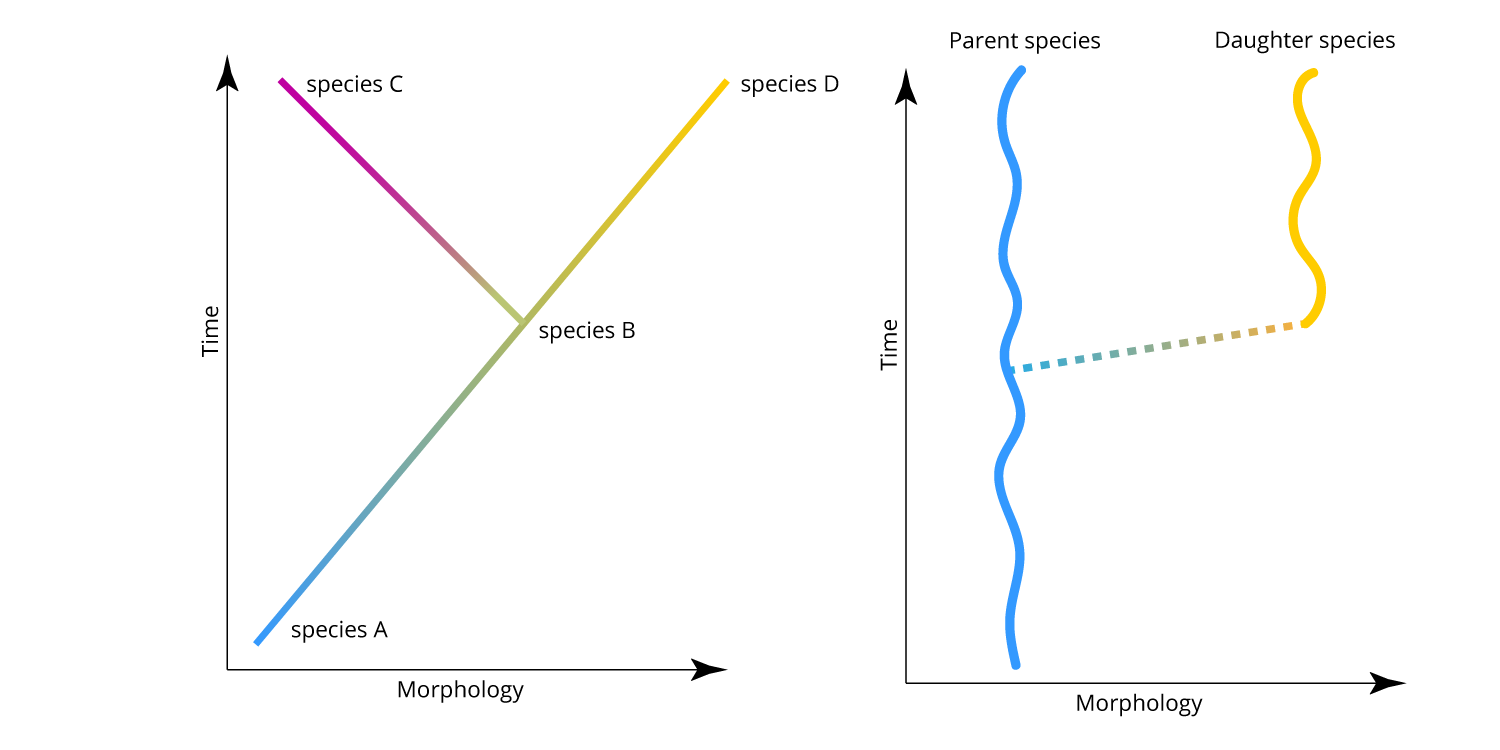
Cladogenesis in the framework of phyletic gradualism (left) and punctuated equilibrium (right). Image by Jonathan R. Hendricks.
The image above shows two frameworks for cladogenesis. The figure on the left demonstrates what Eldredge and Gould called phyletic gradualism: there is lineage splitting, but anagenesis both before and after the splitting event. The figure on the right shows lineage splitting within the framework of punctuated equilibria. An important point to recognize in the punctuated equilibria framework is that the evolution of the daughter species from the parent species does not imply that the parent species goes extinct. This is implied, however, by any model of anagenesis, including phyletic gradualism. Do not trouble yourself too much trying to figure out how anagenesis might work: it does not make any biological sense.
It is clear that cladogenesis has been a critically important force in the history of life on Earth: all of the millions of living species are the descendants of one species that lived billions of years ago. There is simply no way to explain the great diversity of extant bacteria, protists, plants, fungi, and animals in the context of slow, gradual anagenesis. Further, anagenesis does not make much sense in the first place and biologists and paleontologists should never have expected to find significant evidence for it in the fossil record. The reason why speaks to the power of natural selection and the rapidity of evolutionary change that has been demonstrated to occur on human time scales, especially in experimental settings (for numerous examples, see Jonathan Losos' recent book Improbable Destinies).
Just as an overwhelming number of studies that focus on the anatomy of fossil organisms have shown that stasis prevails, an even greater number of studies on modern organisms have shown that the action of natural selection is sufficient in nature to produce substantial changes in morphology over hundreds or thousands of years. This might seem slow and gradual when compared with a human lifespan, but recall that Darwin’s vision of gradualism was put forth in the context of eons of geological time. When it comes to the fossil record, however, a few hundred years, or even a few tens of thousands of years, is a geological instant or "blink of the eye" only rarely likely to be preserved (see DEAL section on the completeness of the fossil record). Moreover, a few thousand years is extremely rapid compared to the millions of years of stasis that characterize many species.

Lucifer Falls at Robert H. Treman State Park, Ithaca, New York (note people for scale). The sequence of rocks shown in this photograph (Devonian Sonyea Group) may represent about 1,000,000 years of geological time. Photograph by Jonathan R. Hendricks.
Because speciation happens so quickly on geological time scales, it is only very rarely expected to be captured in the fossil record. Most depositional settings do not experience continuous sedimentation and new beds may only be laid down sporadically (for example during major storm events). Consider the Devonian rocks that are beautifully exposed at Lucifer Falls near Ithaca, New York (see photograph above). Regionally, these rocks accumulated at a rate of about 1 ft. of thickness every 4,000 years. That means that the sequence of rocks shown in the photograph may span close to 1,000,000 years. Individual beds, however, were quickly deposited in a span of hours. What does this tell us? It suggests that hundreds or thousands of years likely passed between individual depositional events. Thus, most geological time here is "missing time," when the geological tape recorder was not on. It is highly unlikely that speciation could be documented in such an incomplete (if typical) section of rock (speciation could happen over the span of just two or three depositional events). On the other hand, this would be the perfect type of rock section from which to document the long-term morphological stasis of species (and this has been done in rock sections just like this).
Finally, to return to the topic from the previous section regarding the view of species as real and distinct evolutionary lineages, consider that punctuated equilibria is a pervasive pattern: it suggests that species often have a long history, typically spanning millions of years, during which they are independent from other such species. This independent history means that they likely have distinctive features that characterize them and distinguish them from other such species, making them easier for us to identify using both morphological and molecular approaches. The reality and long-term stability of species also could mean that they might be subject to selection, a topic we will discuss later in this DEAL chapter.
References and further reading
Eldredge, N. 1989. Time Frames: The Evolution of Punctuated Equilibria. Princeton University Press, Princeton, NJ.
Eldredge, N. and Stephen Jay Gould. 1972. Punctuated equilibria: an alternative to phyletic gradualism. Pp. 82-115 in: Schopf, Thomas J. M. (ed.), Models in Paleobiology. Freeman, Cooper and Company, San Francisco, CA.
Eldredge, N., and B. S. Lieberman. 2018. Paleontological life lessons, stasis, and the genius of Yogi Berra, a post in the Macroevolutionaries blog, https://bronze-sheep-
29mh.squarespace.com/blog/2018/11/12/asleep-at-the-switch
Eldredge, N., J. Thompson, P. Brakefield, S. Gavrilets, D. Jablonski, J. Jackson, R. Lenski, B. S. Lieberman, M. McPeek, W. Miller, III. 2005. The dynamics of evolutionary stasis. Paleobiology 31:133-145.
Gould, S. J. 2007. Punctuated Equilibrium. Harvard University Press, Cambridge, MA.
Gould, S. J., and N. Eldredge. 1993. Punctuated equilibrium comes of age. Nature 366:223-227.
Hunt, G., M. J. Hopkins, and S. Lidgard. 2015. Simple versus complex models of trait evolution and stasis as a response to environmental change. Proceedings of the National Academy of Sciences, 112: 4885-4890.
Jackson, J. B. C., and A. H. Cheethan. 1994. Phylogeny reconstruction and the tempo of speciation in cheilostome Bryozoa. Paleobiology 20:407-423.
Lieberman, B. S., and N. Eldredge. 2008. Punctuated equilibria. Scholarpedia: 3:3806, http://www.scholarpedia.org/article/Punctuated_equilibria
Lieberman, B. S., C. E. Brett, and N. Eldredge. 1995. Patterns and processes of stasis in two species lineages from the Middle Devonian of New York State. Paleobiology 21:15-27.
Losos, J. B. 2017. Improbable destinies: fate, chance, and the future of evolution. Riverhead Books, 384 pp.
Pagel, M., Venditti, C., and A. Meade. 2006. Large punctuational contribution of speciation to evolutionary divergence at the molecular level. Science 314:119-121.
Stanley, S. M., and X. Yang. 1987. Approximate evolutionary stasis for bivalve morphology over millions of years: a multivariate, multilineage study. Paleobiology 13:113-139.
Content usage
Usage of text and images created for DEAL: Text on this page was written by Bruce S. Lieberman and Jonathan R. Hendricks. Original written content created by Bruce S. Lieberman and Jonathan R. Hendricks for the Digital Encyclopedia of Ancient Life that appears on this page is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Original images created by Jonathan R. Hendricks are also licensed under Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
Content sourced from other websites: Attribution, source webpage, and licensing information or terms of use are indicated for images sourced from other websites in the figure caption below the relevant image. See original sources for further details. Attribution and source webpage are indicated for embedded videos. See original sources for terms of use. Reproduction of an image or video on this page does not imply endorsement by the author, creator, source website, publisher, and/or copyright holder.