

Chapter contents:
Evolution and the Fossil Record
– 1. Natural selection
– 2. Species and species concepts
– 3. Speciation
– 4. Punctuated equilibria and stasis
–– 4.1 Videos about punctuated equilibrium and stasis
– 5. Macroevolution
–– 5.1 Hierarchies
–– 5.2 Species selection ←
–– 5.3 Abiotic vs. biotic causes of macroevolution
–– 5.4 Evolutionary radiations
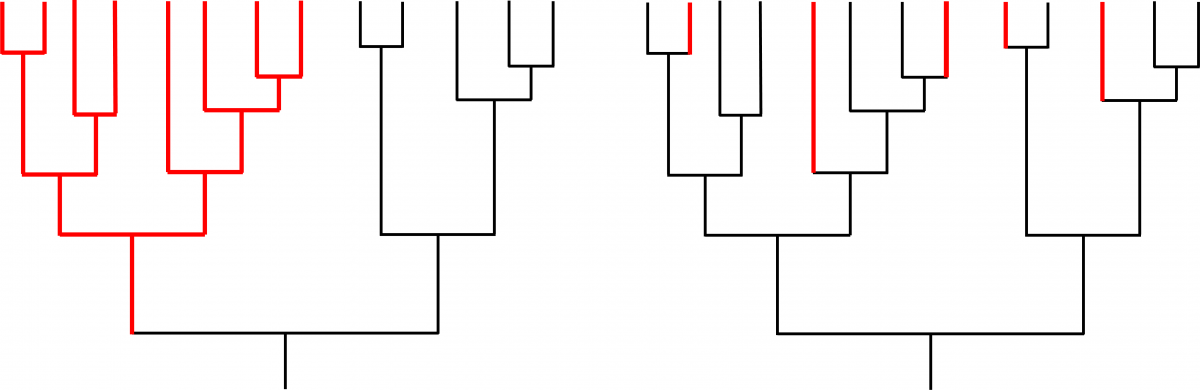
Above image: Two hypothetical examples of diversification in a group of closely related species.
Certain groups (or, clades) of organisms are much more diverse than others. That is to say, they contain more species. As an example, consider the mammals, the group to which we belong. Of the more than 5,000 species of mammals alive today, over 2,000 (~40%) belong to the Order Rodentia (the rodents). Examples of rodents include mice, rats, squirrels, capybaras, and the industrious, dam-building beaver.

American Beaver, by John Audubon (ca. 1844). Part of the collection of the Brooklyn Museum (public domain).
In contrast to the diverse Rodentia, the entire Order Tubulidentata includes only one living species: the aardvark (Orycteropus afer).
"Can You Dig It? meet Zola the Aardvark." Source: San Diego Zoo Kids.
There are many reasons to like the strange aardvark, not least of which is that famous "Late Night" host David Letterman once ranked “Clark the aardvark went to the park after dark and drank Cutty Sark with Marky Mark” as the second coolest thing one could say with a Boston Accent.
Leaving Boston accents aside, a key evolutionary question is: why are rodents so diverse, but aardvarks are represented by just one living species? What is the reason for this great difference in diversity?
One possibility is that the rodent clade is simply much older than the aardvark clade, and therefore has had much more time to diversify. Rodentia originated in the Paleocene, while the oldest known aardvark (which has a much poorer fossil record) is from the Miocene (Lehmann, 2009). Even so, is ~30 million years of geological time enough to account for this great difference in diversity? What other factors might be important? Consider the following possibilities.
1) Rodents might have a higher rate of speciation than aardvarks. This would mean that over a given span of time, speciation would produce more rodent species than aardvark species.
2) Rodents might have a lower rate of extinction than aardvarks. This would mean that over a given span of time, fewer species of rodents would go extinct than aardvarks.
3) A third possibility relates to chance factors. That is to say, rodents might be more diverse than aardvarks simply because of random events in the past that were to the benefit of rodents and/or the detriment of aardvarks.
For now, let's set aside the third possibility (the relationship between randomness and evolution will be discussed in the next chapter). A traditional Darwinian explanation might suggest that rodents are more diverse than aardvarks because they were honed by natural selection to be much more fit to the environments in which they live than aardvarks, and thus are less likely to go extinct. Clearly rodents do something right: we need look no further then the fact that beavers can be highly effective ecosystem engineers, or the fact that rats seem to be inherently tough and capable of surviving in conditions that would be abhorrent to most other mammal species.
"New York City rat taking pizza home on the subway (Pizza Rat)." Source: Matt Little.
There are potential problems, however, with such a traditional Darwinian explanation. For instance, individual organisms within a given species might be very well adapted to the environment in which they live, resulting in an increase in the number of individuals belonging to that species over time. Even so, this proliferation of individuals within the species does not necessarily mean that its speciation rate will increase. Instead, it might lead to a reduction in extinction rate. This is because the more individuals that a species contains, the less likely that it is to go extinct. (We could imagine a situation, however, where a drastic increase in population size could lead to a severe reduction in the amount of available resources needed by the successful species to survive, which ultimately could cause that species to go extinct).
One process that might explain the difference in diversity between various groups (or, clades)--including differences that relate to speciation and extinction rates in individual clades--is species selection. Species selection is an evolutionary process that is logically equivalent to natural selection, but operates at the species level (natural selection operates at the level of individual organisms).
Recall that selection is the interaction between some emergent property of an entity and the surrounding environment that results in the differential survival or differential reproduction of that entity. For example, suppose that some individuals within a particular species of clam have thicker shells than others. If shell crushing predators like crabs are common in the surrounding environment, clam individuals with thicker shells (an emergent property of an individual clam) might be more likely to survive (because they are less commonly crushed) than those that have thinner shells (because they get crushed and are selected against). The spots on a leopard or the speed of an impala (a type of antelope) are also examples of emergent properties of individual organisms, and—like the thickness of a clam's shell—might also provide survival advantages or disadvantages in particular environments, on which selection can operate.

Left: the African leopard, Panthera pardus. Image by JanErkamp (Creative Commons Attribution-Share Alike 3.0 license). Right: the impala, a type of antelope. Image by Hein waschefort (Creative Commons Attribution-Share Alike 3.0 license).
{kind=link}
{kind=link}
The emergent properties of individual organisms (e.g., the thickness of a clam's shell, the pattern of a leopard's spots, or the speed of an antelope) are not the same properties that are emergent at the species level. One of the most obvious properties that is emergent at the species level—but not at the level of individual organisms—is geographic range. Think of your own geographic range: for the most part, you likely spend most of your day-to-day life within a small geographic area (e.g., you might commute across town for work or school, but you usually return to the same house or apartment each night). Collectively, the human species has a much, much larger geographic range than any individual person does (in fact, humans occupy nearly all but the most remote parts of the world).
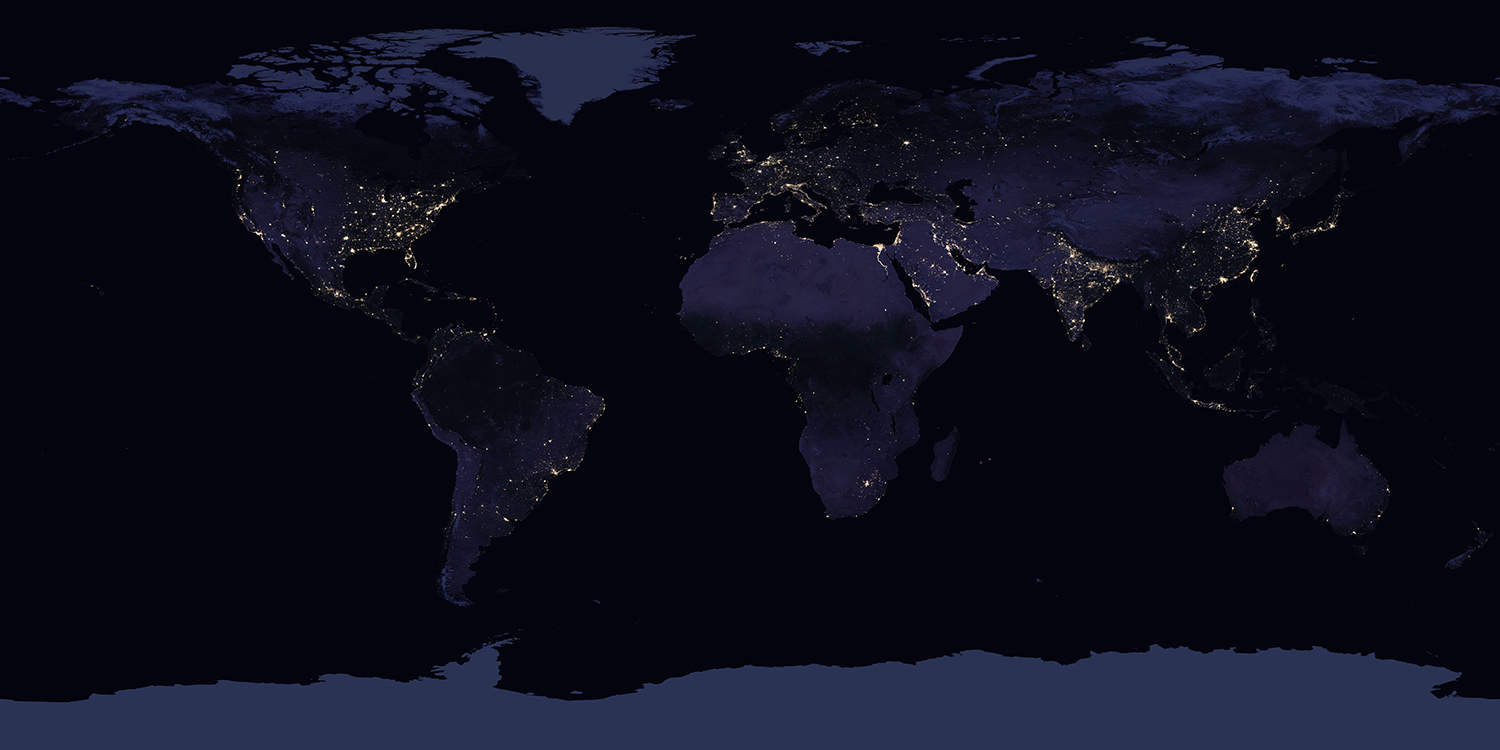
The world at night, lit by human civilization. Humans occupy much of Earth's habitable land area. Source: "Black Marble 2016" by NASA (composite satellite images; public domain).
In this respect, a global geographic range is an emergent property of the human species that is not a sum of our individual parts: it has nothing to do with the morphological variation exhibited by individual people. That range of variation itself is another type of species-level emergent property because individual organisms don't have variation, but species do (e.g., the human species exhibits a range of different heights, but individual humans have a fixed adult height). Similarly, the genetic code of an individual animal is fixed at conception, but there is genetic variation among all individuals within a species. Taking it one step further, some species show greater amounts of genetic variability than others.
Species selection works like natural selection, but selects upon the emergent properties of species, not the emergent properties of individual organisms within species. Species that are selected against may go extinct (equivalent to natural selection selecting against individual organisms with particular disadvantageous variations). Alternatively, species selection may cause some species to speciate at higher rates than others (equivalent to natural selection favoring the reproduction of individual organisms).¹
Consider again the example with which we began: rodents and aardvarks. While species selection has likely never been formally explored as a possible reason why rodents are so much more diverse than aardvarks, it is appropriate to consider it as a possibility. Think about the emergent species-level properties of rodents and aardvarks and generate some hypotheses for why the rodent clade is so much more diverse than the aardvark clade.
¹ Elisabeth Vrba provided a sophisticated and highly logical view of the topic, and it is her framework, along with that of Niles Eldredge, that we are primarily using.
Studies of species selection and gastropods
Most marine gastropods (or, snails) develop in one of two ways. Some species emerge from their eggs on the seafloor and become members of the plankton for several weeks or more. They feed in the plankton and are sometimes pushed great distances by ocean currents before metamorphosing into adults and settling on the seafloor for the rest of their lives. Gastropod species with this mode of development are called planktotrophic (or, planktonic). Juveniles of other gastropod species emerge from their eggs and spend only a couple of days to no time at all in the plankton; some simply emerge from their eggs and crawl away, spending their entire lives on the seafloor. These species have nonplanktotrophic development (more precisely, these species have lecithotrophic or direct development). Because they spend little or no time developing in the plankton, nonplanktotrophic snails spend most of their lives very near the place where they were born. They typically develop in larger eggs than species with planktotrophic development and, because of the greater amount of food available to them early in their development, do not need to feed prior to metamorphosing into adults.
Now consider two closely related snail species, one with planktotrophic development, and one with nonplanktotropic development. Which of the two species do you predict will have a greater geographical range? The answer is pretty easy: the one with planktotrophic development. This is because individual larvae from a single clutch of eggs might become distributed by ocean currents over a broad geographic area before settling on the seafloor and becoming adults. This prediction is supported by direct observations of the geographic ranges of closely related gastropod species with differing developmental modes. For example, compare the distributions of the three species of venomous cone snails shown in the map below. All are members of the same clade, but only one of the species (Conus araneosus) has nonplanktotrophic development. Its range is much smaller than those of Conus marmoreus and C. bandanus, both of which have planktotrophic development.
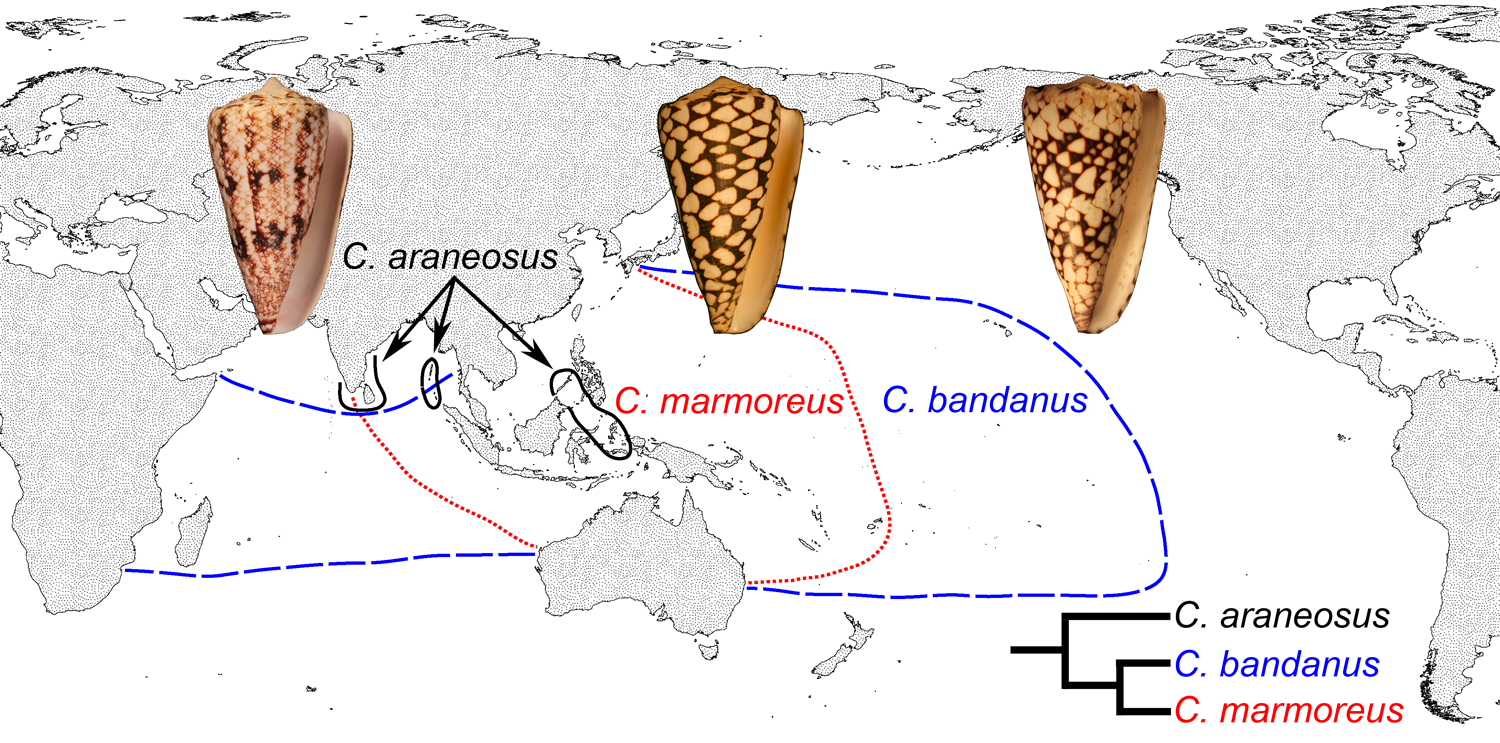
Differing geographical ranges of three closely-related species of cone snails. Conus marmoreus and Conus bandanus have planktotrophic development, while Conus araneosus has nonplanktotrophic development (data from Röckel et al. (1995) and Duda and Kohn (2005)). Image source: Hendricks (2012, fig. 1) in Evolution: Education and Outreach 5:406 (open access).
It has been recognized that those modern gastropod species with nonplanktotrophic larvae typically have higher within-species genetic variation than species with planktotrophic larvae. This has been attributed to the fact that different populations of species with planktotrophic larvae tend to stay in genetic contact with one another because their populations are well connected by their mode of larval dispersal (i.e., populations are continuously "seeded" with new larvae from other populations). This leads to different populations being genetically somewhat similar, despite sometimes being separated by great distances.
By contrast, populations of species with nonplanktotrophic developmental modes are much more likely to become genetically isolated. Given enough time, this isolation is predicted to lead to genetic divergence and eventually speciation. Indeed, on these grounds, it might be expected that species with nonplanktotrophic development should be more likely to speciate than those with planktotrophic development. Alternatively, because they have smaller geographic ranges, nonplanktotrophic species are predicted to be more prone to extinction, as localized environmental perturbations are more likely to wipe out all members of a species. Geographically widespread species may similarly suffer from these events locally, but will maintain healthy populations elsewhere, favoring the geological persistence of such species.
Let's return to thinking about species selection. In the context of marine gastropods, the genetic structure and connectivity that corresponds with each type of developmental mode (as well as the greatly varying geographic range sizes of species with planktotrophic versus nonplanktotrophic development) presents a specific example of a property that is emergent at the species level, but not the level of individual organisms. Having identified this emergent property of gastropods, it then becomes possible to develop hypotheses related to species selection that can be tested, including using the fossil record.
You might be reasonably wondering how it is possible to determine the developmental mode of long-extinct gastropods. As it turns out, the shells of gastropods sometimes reveal their style of larval development. Unless they are eroded or broken away, gastropods retain their larval shells into adulthood at the apex (or, tip) of their shell. This larval shell is often called the "protoconch." Species of gastropods with planktotrophic development usually have protoconchs with three or more whorls. Alternatively, species with nonplanktotrophic development usually have protoconchs with two or fewer whorls. Well-preserved specimens of fossil gastropods often retain their protoconchs, allowing the developmental mode of an individual species to be determined.

Scanning electron photomicrograph (SEM) image of a larval shell of a fossil cone snail shell from the early Pliocene of the Dominican Republic (collections of the Paleontological Research Institution, Ithaca, New York). The arrow indicates the boundary between the protoconch (larval shell) and the subsequent spire whorls. Because the protoconch has more than 3 whorls, this species likely had planktotrophic larval development. This work by Jonathan R. Hendricks and the Digital Atlas of Ancient Life is licensed under a Creative Commons Attribution-Sharealike 4.0 International License.
What does the fossil record tell us about the speciation rates of gastropod species with planktotrophic versus nonplanktotrophic development? What about their relative extinction rates? Thor Hansen (1982) studied the developmental modes of six different families of gastropods from the Paleogene of the U.S. Gulf Coast and found that, over time, the number of species with nonplanktotrophic development increased relative to the number with planktotrophic development. He explained his results by arguing that "higher speciation rates for nonplanktotrophs and a unidirectional trend in developmental change may account for the progressive increase of nonplanktotrophic development" (p. 367). Thus, his results were seemingly consistent with the predictions of species selection.
David Jablonski (1986) further explored differences in macroevolutionary patterns among planktotrophic and nonplanktotrophic Cretaceous gastropods from the U.S Gulf Coastal Plain. He also found that planktotrophic species had much larger median geographic ranges (1,860 km vs. 380 km) than nonplanktotrophic species. As might be predicted, he found that the greater geographic ranges of the planktotrophic species also seemed to confer resistance to extinction: the median geological durations (or "lifespans") of planktotrophic species were longer (6 million years versus 2 million years).
These results found by Hansen and Jablonski are seemingly consistent with the predictions of species selection. There is one shortcoming, however: the reason why the number of nonplanktotrophic species might have increased over time is that planktotrophic lineages may commonly give rise to nonplanktotrophic species. Once a lineage becomes nonplanktotrophic, however, it rarely--if ever--makes the switch back to planktotrophy. This is because the ability to eat food as a larva is lost in nonplanktotrophs and in most cases cannot be re-developed, making a switch back to planktotrophy nearly impossible.
Studies that have analyzed the distribution of planktotrophic and nonplanktotrophic lineages within a phylogenetic context have presented an additional problem. Species selection predicts that lineages with nonplanktotrophic development should show higher rates of speciation than lineages or clades with planktotrophic development (e.g., the phylogeny on the left side of the image below). Instead, studies like those of Lieberman et al. (1993; who studied turritellid snails) and Duda and Palumbi (1999; who studied cone snails) have shown independent origins of nonplanktotrophic species within gastropod clades dominated by species with planktotrophic development (e.g., the phylogeny on the right side of the image below). Thus, the diversity difference documented by Hansen between planktotrophic and nonplanktotrophic species can potentially be explained by the biased conversion of species with planktotrophic development to species with nonplanktotrophic development, with a paucity of reversions to the original state. This alternative could explain the results of Hansen, making it unnecessary to invoke increased speciation rates, or species selection.
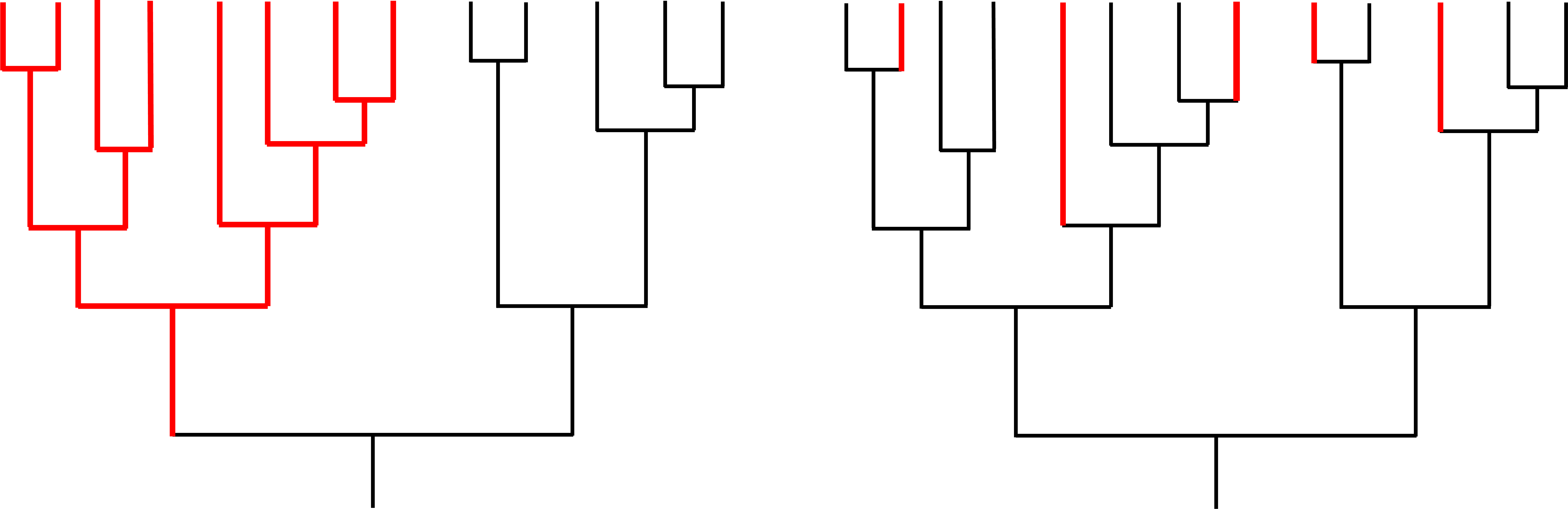
Two hypothetical examples of diversification in a group of closely related species. Black indicates presence of the ancestral condition (e.g., planktotrophic development); red indicates presence of a derived condition (e.g., nonplanktotrophic development). In the example on the left, the origin of the derived condition has caused an increased rate of speciation within a single clade, providing evidence for species selection. In the example on the right, the derived condition has originated multiple times, but there is no evidence that the origin of the trait has increased the speciation rate; therefore, there is not any evidence for species selection. Image by Jonathan R. Hendricks based in part on fig. 1 in Duda and Palumbi (1999).
In the parlance of Elisabeth Vrba, the increase documented by Hansen in the diversity of nonplanktotrophic species relative to planktotrophic species is an effect of processes operating beneath the species level. In this particular case, the pattern could be a result of physical changes emergent at the level of individual organisms (i.e., the "one-way" loss of the ability of nonplanktotrophs to feed during the larval phase), rather than different properties that are emergent at the species level.
This finding doesn’t mean that the population genetic structure seen in species with a nonplanktotrophic developmental mode isn’t a good example of a species-level emergent character. Nor does it meant that there aren’t other potential examples of species selection. It does mean, however, that, although an intriguing idea, there is not a lot of corroboration thus far for the hypothesis that species selection is an important macroevolutionary phenomenon. But it is an idea that merits continued testing and exploration, and belongs in any discussion of evolution in general, and macroevolution in particular.
References and further reading
Duda, T. F., Jr. and A. J. Kohn. 2005. Species-level phylogeography and evolutionary history of the hyperdiverse marine gastropod genus Conus. Molecular Phylogenetics and Evolution 34: 257-272.
Duda, T. F., Jr. and S. R. Palumbi. 1999. Developmental shifts and species selection in gastropods. Proceedings of the National Academy of Sciences, USA 96: 10272-10277.
Gould, S. J. 2002. The structure of evolutionary theory. Cambridge: Harvard University Press.
Hansen, T. A. 1982. Modes of larval development in Early Tertiary neogastropods. Paleobiology 8(4): 367-377.
Hendricks, J. R. 2012. Using marine snails to teach biogeography and macroevolution: the role of larvae and dispersal ability in the evolution and persistence of species. Evolution and Outreach 5: 406.
Jablonski, D. 1986. Larval ecology and macroevolution in marine invertebrates. Bulletin of Marine Science 39(2): 565-587.
Lehmann, T. 2009. Phylogeny and systematics of the Orycteropodidae (Mammalia, Tubulidentata). Zoological Journal of the Linnean Society 115: 649-702.
Lieberman, B. S. and E. S. Vrba. 1995. Hierarchy theory, selection, and sorting. BioScience 45(6): 394-399.
Lieberman, B. S. and E. S. Vrba. 2005. Stephen Jay Gould on species selection: 30 years of insight. Paleobiology 31(2, supplement): 113-121.
Röckel, D., W. Korn, and A. J. Kohn. 1995. Manual of the living Conidae. Grillparzerstr: Verlag Christa Hemmen.
Stanley, S. M. 1975. A theory of evolution above the species level. Proceedings of the National Academy of Sciences, USA 72: 646-650.
Vrba, E. S. 1984. What is species selection? Systematic Zoology 33: 318-328.
Vrba, E. S. and N. Eldredge. 1984. Individuals, hierarchies and processes: towards a more complete evolutionary theory. Paleobiology 10: 146-171.
Content usage
Usage of text and images created for DEAL: Text on this page was written by Bruce S. Lieberman and Jonathan R. Hendricks. Original written content created by Bruce S. Lieberman and Jonathan R. Hendricks for the Digital Encyclopedia of Ancient Life that appears on this page is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Original images created by Jonathan R. Hendricks are also licensed under Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
Content sourced from other websites: Attribution, source webpage, and licensing information or terms of use are indicated for images sourced from other websites in the figure caption below the relevant image. See original sources for further details. Attribution and source webpage are indicated for embedded videos. See original sources for terms of use. Reproduction of an image or video on this page does not imply endorsement by the author, creator, source website, publisher, and/or copyright holder.