


Chapter contents:
Evolution and the Fossil Record
– 1. Natural selection
– 2. Species and species concepts ←
– 3. Speciation
– 4. Punctuated equilibria and stasis
–– 4.1 Videos about punctuated equilibrium and stasis
– 5. Macroevolution
–– 5.1 Hierarchies
–– 5.2 Species selection
–– 5.3 Abiotic vs. biotic causes of macroevolution
–– 5.4 Evolutionary radiations
Above image: A very small sampling of some of the millions of different species from Earth's past and present.
Life's hierarchies
One of the most fundamental features of the natural world is that there are hierarchies of relationship that connect all living things.
To the best of our knowledge, all life on Earth today shares a single common ancestor that lived billions of years ago. Every organism that we know of--whether alive today or that died hundreds of millions of years ago--descended from this ancestor. Further, there is a line of relationship--of parents and their offspring--that connect you to this ancient shared common ancestor. There are also, however, many side branches that are not part of your direct ancestry.
Think about your own family tree: you descended directly from your grandparents via your mother, but your grandparents also may have had other children besides your mother (that is, your aunts or uncles), who in turn had children of their own (your cousins).
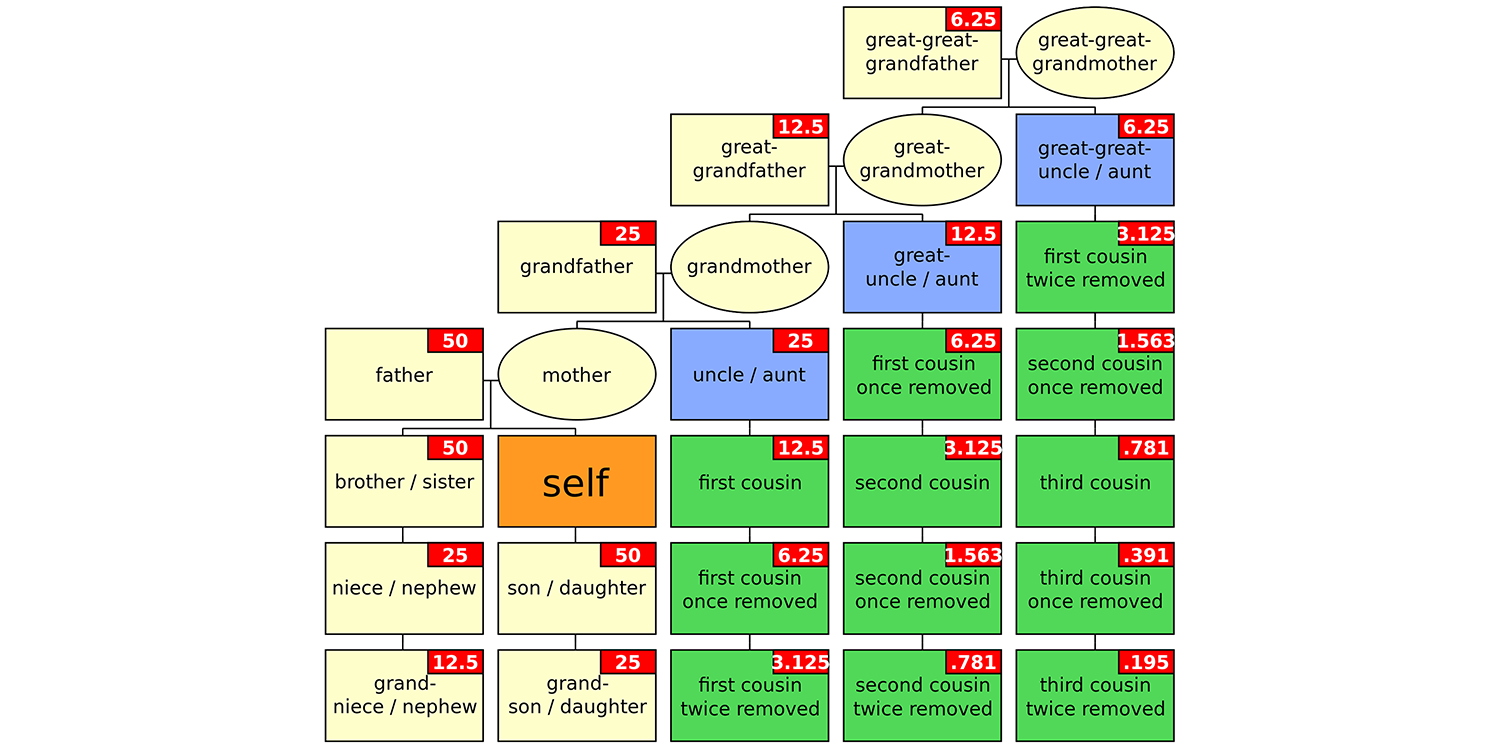
Hierarchy of familial relationships. Numbers in red boxes indicate percentages of genetic relationship relative to yourself. Image source: "Gringer" (Wikimedia Commons; public domain).
{kind=link}
The same is true when considering evolutionary patterns at larger scales. Evolution has produced a bush of branches, none leading to one type of organism in particular (just as your own family tree does not lead to any particular "most important" person (contrary to what your cousins might try to tell you over Thanksgiving dinner!). Because of this, evolution must be thought of as hierarchical, just as a family tree is hierarchical. We call evolution's family tree the "tree of life."

An artistic representation of the Tree of Life. Source: the Tree of Life Web Project (Creative Commons Attribution-No Derivative Works License - Version 3.0).
Source: "Tree of Life video HD, narrated by David Attenborough (Youtube).
Another fundamental aspect of evolution and the history of life is that biological diversity exists in discontinuous packages. Evolutionary history has produced numerous branches, each composed of groups of organisms that are more similar to each other than they are to groups on other branches. Ants look like ants, hawks look like hawks, and frogs look like frogs. If you carefully study their bodies, however, you will find that frogs and hawks share much more in common with each other than they do with ants (most obviously, they both have bones, which ants lack). Within any of these broad groups, it is possible to define subgroups more-and-more narrowly until one is presented with very similar clusters of individual organisms. We call these clusters species, and more than 1.2 million of them have been described by scientists thus far. Systematists (scientists who study biological diversity, or biodiversity, and the relationships among organisms) estimate, however, that there may be between 10 and 30 million species alive on Earth today. This estimate does not include the number of species known only from the fossil record.

A very small sampling of some of the millions of different species from Earth's past and present. Image by Jonathan R. Hendricks.
The Systematics chapter of the Digital Encyclopedia of Ancient Life covers the practice of assigning scientific names to species and higher level groupings. It also introduces phylogenetic trees, which are branching diagrams that depict the degrees of relationship among species or higher taxonomic groups (that is, they represent certain portions of the tree of life).

An example of a phylogeny, in this case showing the relationships among most major groups of life. The small "You are here" indicator in the upper left corner of the phylogeny indicates the position of Homo sapiens (you and me). Source: David Hillis lab.
For now, though, let's simply concern ourselves with the nature of species as biological entities. Their existence is clearly a phenomenon that evolution needs to explain. Their characterization and definition, however, has long proven vexing, even if each of us has our own "gut feeling" for what they represent.
What are species?
For a long time the notion of what species are and what they represent has often been treated as one of the great philosophical mysteries, along with whether gravitational waves exist...
Source: "Gravitational waves hit the Late Show" on The Late Show with Stephen Colbert (Youtube).
whether life exists elsewhere in the universe...
Source: "Drake equation visualized by Carl Sagan" on the program Cosmos (Youtube).
and how might history have turned out differently if Rose had made a little room for Jack on that floating door at the end of Titanic?
Source: "A "Titanic" Myth: Would jack Have Survived if Rose Had Shared the Door?" by Mythbusters on Science Channel (Youtube).
Any discussion of species presents two importation questions:
- What "are" species as evolutionary entities? (Or, what are species in nature?); and
- How should we recognize and define species? (Or, what are species in practice?).
Nearly all of the over one million described living species of plants, animals, and other lifeforms have been named on the basis of their morphological appearances (or, how they look). But, species are groups of organisms that not only resemble each other closely in terms of their anatomy. They also seem to comprise individuals of both sexes that will at times reproduce with one another. We can very easily go out into the natural world and identify these minimal clusters of individuals consisting of at least one male and one female, defining them by one or more distinct morphological features that they--and only they--possess.
Consider the American robin, or Turdus migratorius (a rather crude-sounding name, if you think about it). This common bird is a harbinger of spring for many Americans who live in cold, northern climates.
A short video about the American robin. Source: "The American Robin" by LesleytheBirdNerd.
The American robin occurs (at least during certain times of the year) from Mexico all the way to the Northwest Territories of Canada.
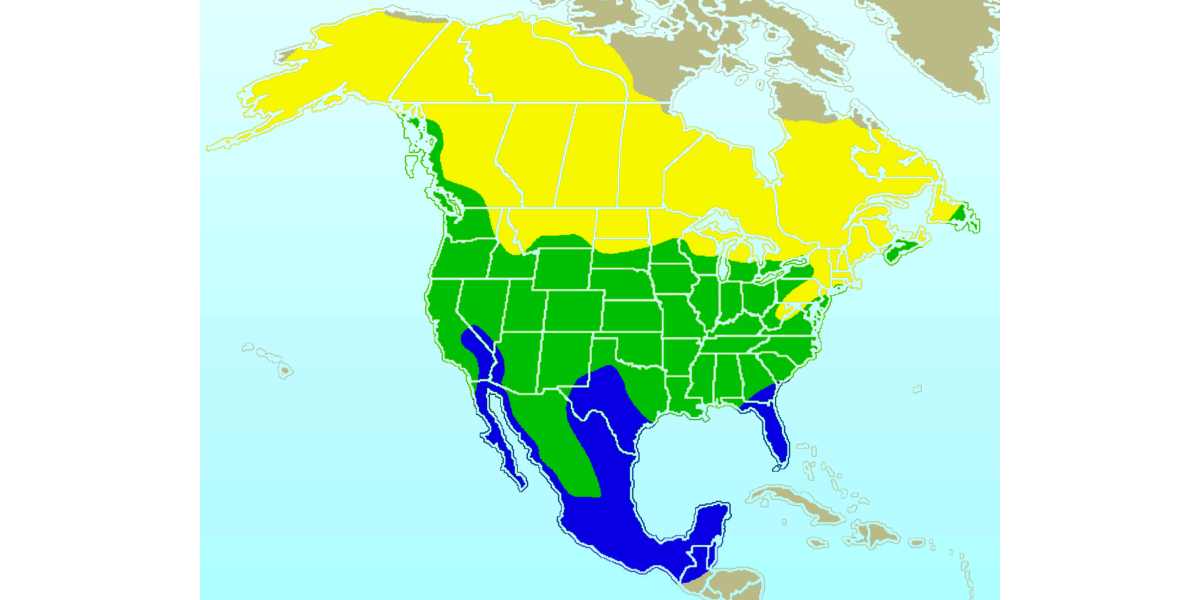
Range map of the American robin. Yellow indicates the robin's breeding range; green, its year-round range; and blue, its wintering range. Source: Image by Ken Thomas (public domain).
{kind=link}
Throughout their range, individual American robins always posses the same basic appearances and types of behaviors. We suppose, based on the scientific name of their genus (i.e., Turdus) that one of those behavioral proclivities includes pooping on freshly washed cars. (Just kidding! Turdus is actually the Latin name for a thrush, the broader grouping of birds to which robins belong). Another notable behavior of robins is their odd, though distinctive song.
Video demonstrating the call of the American Robin. Source: "Robin Call" by Corey Schmaltz, TheBackyardBirder.
Within this very large cluster of individual birds we’d have a hard time using morphological features to recognize narrower groupings. American robins all more-or-less look alike.
We call such similar groupings of individual organisms a species. The existence of things like species of robins, or trilobites, or mushrooms, or flowers is one of the manifest patterns in nature and it also is therefore probably telling us something significant about the way the evolutionary process works.

A trilobite (Paradoxides), mushroom (fly agaric, Amanita muscaria), and flower (California poppy, Eschscholzia californica). Mushroom image source: Michael Maggs (Wikimedia Commons; Creative Commons Attribution-Share Alike 2.5 Generic license). Other images: Jonathan R. Hendricks.
.JPG){kind=link}
How are species defined?
Biologists have developed numerous technical definitions of species. Most students learn only the "biological species concept" in their introductory life science courses. This concept, championed by ornithologist Ernst Mayr, argued that species "are groups of actually or potentially interbreeding natural populations which are reproductively isolated from other such groups." Under this definition, a mule (the infertile offspring of a male donkey and a female horse) is not a species because it cannot reproduce with other mules.

A pack mule, seemingly oblivious to the fact that it is not a species. Source: image by Dario Urruty (public domain via Wikimedia Commons).
{kind=link}
The biological species concept, however, presents some practical challenges. For example, to "prove" that two individuals (a male and female) belong to the same species, one would need to confirm that they mate in nature, then go on to produce offspring that are also fertile. This would be next to impossible to study in many habitats (e.g., for species that live on the bottom of the ocean).
The "morphological species concept" is somewhat more practical. It defines species as groups of individuals that are morphologically similar to one another and are morphologically distinct from other such groups. Another way of saying this is that members of the same species look alike; members of different species look different. A related variant of the morphological species concept is the "phylogenetic species concept," which defines species as "the smallest aggregation of (sexual) populations or (asexual) lineages diagnosable by a unique combination of character states" (Wheeler and Platnick, 2000, p. 58).
It turns out that the morphological and phylogenetic species concepts are not ideal, however, because they don't really define what species represent in an evolutionary context. Indeed, species are not just a collection of carefully defined morphological characters; their features are just the information that we use to identify them. This is similar to how bar codes identify foods at the grocery store checkout line, but do not represent any meaningful qualities about the foods themselves (e.g., appearance, taste, odor, etc.).
There are many, many other species concepts (as suggested by the image below). We could go into great detail about these, but we won’t do that because many of them aren’t particularly relevant, or even necessarily accurate.

A handful of the species concepts (and reflections on species) that adorn the ceiling of the paleontology graduate student office in Snee Hall at Cornell University. Images provided by Caren Shin.
Instead, we are going to focus on the one species concept that we think really matters: the "evolutionary species concept." While originally conceived by paleontologist George Gaylord Simpson, our modern understanding of this species concept derives mostly from retired University of Kansas biology professor Ed Wiley.

University of Kansas Curator Emeritus of Ichthyology, Edward Wiley.
Before getting into the particulars of the Evolutionary Species Concept, let's consider some generalities. First, you have already learned about the Biological Species Concept, which stresses the ability of organisms to reproduce (or not) as being central to their reality in nature. We do think that this is true (at least for many sexually reproducing organisms). The ability of some similar organisms to reproduce with each other, but not members of other groups, gives them cohesion and distinctiveness. Consider that if there are two different “looking” groups that are continually interbreeding with each other, then eventually they will become a single, blended group that combines features of both groups: they won’t look different any more. We would only expect the two groups to stay distinct and persist over geological time scales if they are not reproducing with each other. This element of geological time is a key component of the Evolutionary Species Concept.
Wiley and colleague Richard Mayden characterized the Evolutionary Species Concept thusly in 2000: "An evolutionary species is an entity composed of organisms that maintains its identity from other such entities through time and over space and that has its own independent evolutionary fate and historical tendencies" (p. 73).
While this is a very technical definition, note each important attribute:
- "an entity composed of organisms that maintains its identity from other such entities"—members of the same species interbreed with one another, thus maintaining their genetic distinctiveness relative to other species; however, organisms belonging to separate species do not interbreed with each other (this is similar to the Biological Species Concept).
- "through time and over space"—species have their own unique temporal (in some cases spanning millions of years) and geographic histories (note that both time and geography are missing from the Biological Species Concept).
- "independent evolutionary fate and historical tendencies"—the histories of closely related species are independent and the fate (e.g., extinction) of one does not have anything to do with the fate of the other(s).
Species as hypotheses
The Evolutionary Species Concept captures what we think species "are" in nature. Like the Biological Species Concept, the Evolutionary Species Concept is not practical for actually going outside and identifying species (whether extant or from the fossil record). Specialists on different types of organisms use very different tools, approaches, and criteria for recognizing species. For example, genetic data are increasingly being used to differentiate some modern species and are required for recognizing different species of morphologically simplistic organisms such as bacteria. Nearly all named species of plants and animals (both extant and long extinct), however, were named on the basis of their morphology alone. For instance, most species of clams and snails were described only from their shells (essentially just one organ system).
The fundamental problem for recognizing species is the issue of differentiating intraspecific variation (=within species variation) from interspecific variation (=variation between species). (Note: remember "intramural sports" when you think of the prefix "intra" - they are competitions between groups at a single college or university; remember "intercollegiate sports" when you think of the prefex "inter" - they are competitions between different colleges or universities.) The intra- and interspecific variation that one is most often faced with is morphological—and this is always the case, of course, when considering species in the fossil record—but may also be genetic, or even behavioral.

Beetle specimens on display at the Melbourne Museum. How many species do you think are represented? Do any of the specimens belong to the same species? Photograph by Jonathan R. Hendricks.
How much variation is enough to differentiate two similar species? If two groups overlap in a particular range of measurements, should they be considered different species? What about if they vary genetically by a tenth of a percent? How much variation is enough variation to recognize two (or more) species? Unfortunately, there are not universal answers to these questions. In fact, these issues sometimes divide taxonomists into two separate camps, lumpers and splitters. Lumpers are taxonomists who assume that species show lots of natural variation. Splitters define species narrowly and do not accept much variability when they circumscribe species. Which view is correct?
At the end of the day, species—as a practical matter—should be thought of as hypotheses to be tested in the face of evidence. Regardless of what species "are", their recognition in nature is a business subject to the biases, preconceptions, and data available to the biologists and paleontologists who study them. Special care must be taken to describe new species (the taxonomic rules for how species are named are described in the Systematics chapter). Retired University of Washington biologist Alan Kohn perhaps put it best: "absolutely no honor is associated with describing a new species. There is only responsibility — a heavy responsibility, for the author of a new species stakes his reputation on his defense of the new name as denoting a real, previously undescribed species, representing breeding populations of many individuals, outside the range of variation of all previously described species" (Kohn, 1980, p. 7; bold emphasis in original).

The extinct cone snail Conus adversarius. The species is unique among cone snails in having a shell that opens to the left (sinistral coiling); all others have shells that open to the right (dextral coiling). Note the subtle morphological differences between the individual specimens. Source: Jonathan R. Hendricks and the Neogene Atlas of Ancient Life (Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License).
Even once a new species is formally named and described, it remains a hypothesis. New specimens might be discovered, for instance, that show features that were not preserved in the originally described specimens. Such additional information could potentially reveal the presence of two species, rather than just one (thereby rejecting the hypothesis of a single species). Alternatively, the discovery of additional specimens might instead reinforce the original description, adding strength to the hypothesis of there being just one species.
Just as all scientific hypotheses are best supported by multiple lines of evidence, hypotheses of species boundaries are similarly strengthened by the presence of multiple lines of evidence. This is why it is generally poor practice to describe new species on the basis of single specimens, especially when they are fragmentary. Such specimens cannot demonstrate natural variation and may often represent extreme morphologies of previously described species.
One early and paleontologically sigificant example of using multiple lines of evidence to define species comes from research on a group of marine invertebrate animals called bryozoans (or, "moss animals"). Bryozoans are colonial animals that live in modern marine and freshwater environments. They also have a diverse fossil record.
Interactive 3D model of a fossil bryozoan (Polypora elliptica) from the Pennsylvanian Thrifty Formation of Texas (PRI 76722). This specimen is from the collections of the Paleontological Research Institution, Ithaca, New York.
Paleontologists Jeremy Jackson and Alan Cheetham were interested in learning whether ancient bryozoan species that were defined only on the basis of their morphology ("morphospecies") were equivalent to modern bryozoan species. To resolve this problem, they studied the morphology and genetics of modern species and found that these "morphospecies are genetically distinct" and that their morphological boundaries are heritable. This was good news, as it meant that species known only from the fossil record were directly comparable as units of analysis to modern species that are genetically unique.
The study of evolution and the description of life's biodiversity both benefit from such multi-pronged approaches, which have been given the name "integrative taxonomy." Much research is now underway to describe (or re-describe) biodiversity using as many lines of evidence as possible. This is a great example of scientific hypothesis testing at work in systematic biology.
References and further reading
Allmon, W. D., and M. M. Yacobucci (eds.). 2016. Species and speciation in the fossil record. University of Chicago Press, Chicago, 384 pp.
Brooks, D. R., and D. A. McLennan. 2002. The nature of diversity: an evolutionary voyage of discovery. University of Chicago Press, Chicago, 676 pp.
Dayrat, B. 2005. Towards integrative taxonomy. Biological Journal of the Linnean Society 85(3): 407-417.
Eldredge, N. 1985. Unfinished Synthesis. Oxford University Press, New York, 248 pp.
Futuyma, D. J., and M. Kirkpatrick. 2013. Evolution, 4th edition. Oxford University Press, New York, 594 pp.
Hendricks, J. R., E. E. Saupe, C. E. Myers, E. J. Hermsen, and W. D. Allmon. 2014. The generification of the fossil record. Paleobiology 40: 511-529.
Jackson, J.B.C., and A. H. Cheetham. 1990. Evolutionary significance of morphospecies: a test with cheilostome Bryozoa. Science 248: 579-583.
Kohn, A. J. 1980. Conus descriptions aren't improving. Hawaiian Shell News 28(5): 7-8.
Mayr, E. 1982. The Growth of Biological Thought: Diversity, Evolution, and Inheritance. Harvard University Press, Cambridge, MA, 996 p.
Padial, J. M., A. Miralles, I. de la Riva, and M. Vences. 2010. The integrative future of taxonomy. Frontiers in Zoology 7:16.
The Tree of Life web project. Website: http://tolweb.org/tree/phylogeny.html.
Wheeler, Q. D., and R. Meier (eds.). 2000. Species Concepts and Phylogenetic Theory: A Debate. Columbia University Press, NY, 230 p.
Wheeler, Q. D., and N. I. Platnick. 2000. The phylogenetic species concept (sensu Wheeler and Platnick). Pp. 55-69 in Wheeler, Q. D., and R. Meier (eds.), Species Concepts and Phylogenetic Theory: A Debate. Columbia University Press, NY, 230 p.
Wiley, E. O., and B. S. Lieberman. 2011. Phylogenetics, 2nd edition. J. Wiley & Sons, New York, 432 p.
Wiley, E. O., and R. L. Mayden. 2000. The evolutionary species concept. Pp. 70-89 in Wheeler, Q. D., and R. Meier (eds.), Species Concepts and Phylogenetic Theory: A Debate. Columbia University Press, NY, 230 p.
Content usage
Usage of text and images created for DEAL: Text on this page was written by Bruce S. Lieberman and Jonathan R. Hendricks. Original written content created by Bruce S. Lieberman and Jonathan R. Hendricks for the Digital Encyclopedia of Ancient Life that appears on this page is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Original images created by Jonathan R. Hendricks are also licensed under Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
Content sourced from other websites: Attribution, source webpage, and licensing information or terms of use are indicated for images sourced from other websites in the figure caption below the relevant image. See original sources for further details. Attribution and source webpage are indicated for embedded videos. See original sources for terms of use. Reproduction of an image or video on this page does not imply endorsement by the author, creator, source website, publisher, and/or copyright holder.